
Os neurocientistas estudam todos os elementos do sistema nervoso com o intuito de compreender como ele está organizado, como funciona, como se forma e como se recupera de lesões.
Imagem: Shutterstock.com
Neurociência aplicada ao estudo da aprendizagem e recuperação motora.
Conhecer os conceitos básicos sobre Neurociência aplicada à aprendizagem e recuperação dos movimentos após lesões neurológicas para que seja possível obter um adequado raciocínio clínico e para que os processos diagnóstico e terapêutico possam contribuir diretamente para a sua atuação profissional.
Identificar os campos de estudo em neurociência e os conceitos básicos sobre neuroimagem funcional
Diferenciar os processos de aprendizagem motora e memória e aprendizagem associativa e não associativa
Descrever os mecanismos cerebrais relacionados à recuperação de lesões neurológicas
A Neurociência é uma ciência multidisciplinar que estuda o sistema nervoso e é uma das últimas grandes fronteiras do conhecimento.
A palavra “Neurociência” deriva de "neurônio", cujo significado é "nervo", mais a palavra "ciência".
Para melhor compreender como esse complexo órgão funciona, existem muitas áreas dentro da Neurociência que abrangem aspectos celulares, evolutivos, computacionais, moleculares, funcionais, comportamentais e médicos do sistema nervoso.
A Neurociência é um campo de estudo que integra, por exemplo, Matemática, Física, Linguística, Engenharia, Ciência da Computação, Química, Filosofia, Biologia, Psicologia, Educação Física, Medicina etc.
Alguns dos exemplos de áreas estudadas na Neurociência são a sinalização neuronal e padrões de conectividade axonal, desenvolvimento neuronal e função biológica, funcionamento cognitivo, neuroimagem relacionada ao diagnóstico de doenças, entre muitas outras. A motivação por trás dessas áreas é que haja melhor compreensão dessas relações para fornecer percepções mecanicistas sobre como o cérebro vincula fatores genéticos, biológicos, psicológicos, comportamentais, sociais e ambientais multiníveis com a saúde física, especialmente vulnerabilidade e resiliência contra outras condições clínicas.
Identificar os campos de estudo em neurociência e os conceitos básicos sobre neuroimagem funcional
O termo “Neurociência” é relativamente novo, entretanto, o estudo do encéfalo é tão antigo quanto a própria ciência. Historicamente, os neurocientistas que se voltaram à compreensão do sistema nervoso vieram de diferentes disciplinas científicas: Medicina, Biologia, Psicologia, Física, Química e até da Matemática.
O salto nas neurociências ocorreu quando esses cientistas perceberam que a melhor abordagem para uma compreensão de como funciona o encéfalo vinha de um enfoque interdisciplinar, ou seja, da combinação de abordagens tradicionais visando a produzir uma nova síntese, uma nova perspectiva.
A maioria das pessoas envolvidas na investigação científica do sistema nervoso considera-se, atualmente, neurocientista.
Compreender como funciona o encéfalo requer conhecimentos sobre muitas coisas, desde a estrutura da molécula até as propriedades eletroquímicas do encéfalo para poder entender, por exemplo, a razão pela qual o cão de Pavlov salivava quando uma campainha tocava no seu clássico experimento sobre condicionamento.
Trata-se de uma experiência revolucionária que ocorreu há 120 anos e demonstrou como funciona o condicionamento de animais. Ivan Pavlov, médico russo do início do século XX, treinou cachorros para que eles ficassem com água na boca sem que houvesse nenhuma comida por perto. Toda vez que os bichos eram alimentados, o médico tocava uma sineta. Com o tempo, os cães começaram a associar as badaladas à comida e chegavam a babar famintos só de ouvir o sino, mesmo que o prato deles estivesse vazio. Muitos podem lembrar que já ensinaram truques parecidos para seus cãezinhos, mas a experiência de Pavlov tinha um propósito bem mais nobre do que disciplinar o melhor amigo do homem. A ideia era propor uma novidade científica: os reflexos condicionados.

O cérebro é uma notável peça de maquinário biológico que fascina os cientistas e o público em geral, enquanto testa os limites de nossa capacidade de compreender nossas próprias mentes.
Cada faceta de função neural requer orquestração intrincada e contínua afinação e remodelação. Os circuitos neurais exibem uma adicional camada de integração que transcende a complexidade de qualquer célula, com características dinâmicas, cuja análise requer computação sofisticada.
Não é surpreendente que essa máquina possa funcionar mal de inúmeras maneiras, levando à debilitação e distúrbios, às vezes, devastadores ao longo da vida.
Assim, compreender o cérebro é intrinsecamente fascinante e altamente relevante para o bem-estar de humanos, além de ser considerado, frequentemente, como o maior desafio nas biociências, possivelmente em toda a ciência moderna.
Consequentemente, nas últimas duas décadas, o campo da Neurociência passou por uma revolução silenciosa, redefinindo suas fronteiras além das ciências biomédicas para incorporar conhecimento e ferramentas de Física, Matemática e Engenharia, assim como as Ciências Sociais e Humanas.
A crescente integração entre a Neurociência e outros campos da ciência vem gerando um grande impacto para sua aplicação. A influência da pesquisa de Neurociência básica em Biologia e Medicina continua a aumentar à medida que os avanços em muitas áreas estão sendo traduzidos em abordagens terapêuticas, incluindo melhores ferramentas para estudar:
Estrutura e função do cérebro na saúde e na doença.
Análise molecular de receptores, canais iônicos e amplas vias moleculares.
Abordagens poderosas para examinar e manipular circuitos neurais.
Estratégias eletrofisiológicas mais sofisticadas para monitorar e modificar a função neural.
Além disso, há uma necessidade cada vez maior de comunicar informações sobre neurociências em todos os níveis, desde a edição de periódicos acadêmicos até a educação do público em geral. Em nível nacional, há também uma grande necessidade de informar reguladores e formuladores de políticas.
O crescimento da área, junto com a comercialização de novos produtos e serviços, resultará na ampliação das oportunidades de carreira nos setores público e privado, incluindo cientistas de bancada, empresários, analistas, consultores e especialistas em propriedade intelectual.
Dado que o número de neurocientistas aumentou muito mais rapidamente do que em qualquer outro campo da pesquisa biomédica nos Estados Unidos, um desafio que o mercado deve enfrentar se refere aos programas de treinamento.
Será que estão atendendo às necessidades atuais e futuras da força de trabalho da área?
Claramente, é preciso atrair os melhores e os mais brilhantes de diversas origens para assumir a tarefa de compreender o cérebro. O desafio atualmente é o de como treinar e reter uma força de trabalho talentosa para garantir um futuro brilhante para a neurociência em geral e para os jovens cientistas que estão entrando na área (BEAR et al., 2008).
Técnicas de neuroimagem não invasivas têm sido de crescente interesse para examinar as mudanças comportamentais e os mecanismos neurais, resultando no desenvolvimento de novas abordagens para melhorar a reabilitação motora.
As técnicas de neuroimagem foram consideradas como promissoras para avaliar a conectividade cortical em função de diferentes tipos de tratamentos, como medicamentos, técnicas de reabilitação física e cognitiva, exercício físico, compreendendo as variáveis neurofisiológicas relacionadas com a aplicação dessas técnicas e, finalmente, para fornecer uma melhor compreensão quanto aos seus benefícios.
Ferramentas não invasivas de pesquisa do cérebro, como a Tomografia por Emissão de Pósitrons (TEP) e a Ressonância Magnética funcional (RMf), têm sido usadas para examinar as mudanças nos mecanismos neurais durante os tratamentos em pacientes com lesões neurológicas.

Tomografia por Emissão de Pósitrons (TEP)

Ressonância Magnética funcional (RMf)
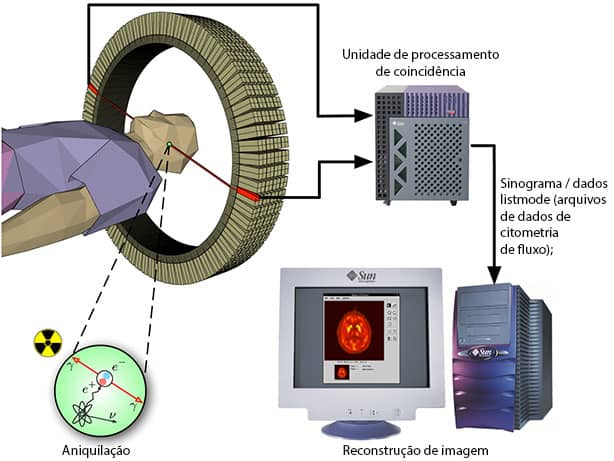
As técnicas de mapeamento cerebral são vitais para a compreensão dos mecanismos moleculares, celulares e funcionais. Dentre eles a Tomografia por Emissão de Pósitrons (TEP) é uma ferramenta extremamente eficaz para o mapeamento da organização funcional no cérebro humano. Essa técnica produz uma imagem tridimensional de processos funcionais no organismo.
Para realizar uma varredura, um isótopo traçador radioativo curto é injetado no paciente (geralmente na corrente sanguínea). Esse traçador é quimicamente incorporado a uma biologicamente molécula ativa, normalmente uma substância como a glicose, que pode ser metabolizada pelas células do corpo.
A TEP produz principalmente informações funcionais e fisiológicas e as estruturas anatômicas são, por sua vez, difíceis de identificar. Portanto, a TEP geralmente deve ser combinada com métodos anatômicos, como tomografia computadorizada ou imagem por ressonância magnética.
A Ressonância Magnética (RM) é uma técnica que permite a visualização de imagens tomográficas tridimensionais de alta resolução sem radiação ionizante. Essa tecnologia também tem permitido a avaliação de características patológicas de forma não invasiva. Além disso, é amplamente utilizada para estudar a organização funcional (Ressonância Magnética funcional – RMf) do cérebro humano, em que os registros de sinais neuronais são apresentados em resposta a estímulos magnéticos.
A RMf não é voltada especificamente para investigar aspectos clínicos, mas também é usada para estudar indivíduos saudáveis e mudanças funcionais em seus cérebros.
A análise de RMf é baseada em uma tecnologia que usa um forte campo magnético e ondas de rádio para capturar alterações hemodinâmicas e metabólicas induzidas por atividade neural, ou seja, por aumentos locais no volume sanguíneo, fluxo e oxigenação.
Esse aumento ativa um intenso campo magnético resultante do alinhamento de suas spins.
Orientação das partículas subatômicas quando imersas em um campo magnético.
Então, quando o pulso de radiofrequência é lançado na representação de sangue, os giros se movem de sua orientação original a um estado de energia excitado.
Durante esse processo, as spins nucleares tendem a retornar para sua condição inicial, ou seja, o estado de menor energia. Porém, quando isso ocorre, o excesso de energia é emitido na forma de radiação eletromagnética.
Essa energia é detectada pelo dispositivo RMf e permite a formação de imagens anatômicas.
RMf tem sido uma ferramenta amplamente usada no mapeamento cerebral para pesquisa, pois permite o estudo de redes neurais de forma dinâmica, rastreando respostas específicas em várias escalas espaciais. No entanto, a RMf pode medir apenas alterações hemodinâmicas, como variações no volume de sangue, ou susceptibilidade magnética intravascular. Além disso, independentemente de ser uma técnica incipiente, a RMf foi aplicada a inúmeros estudos funcionais, variando de experimentos simples a complexos, como investigações neuropsicológicas.
Assim, destaca-se por permitir a exploração de várias funções cerebrais em virtude da sua alta capacidade de diferenciar tecidos.
Existem vários métodos adicionais que podem ser usados para estudar a contribuição de redes corticais específicas para funções cognitivas e motoras. Por exemplo, TEP e RMf são capazes de revelar redes cerebrais envolvidas em funções específicas, no entanto, essas técnicas têm resolução temporal e não podem demonstrar se uma área cortical particular é essencial para uma função específica.
O especialista Sergio Eduardo de Carvalho Machado justifica o uso da neuroimagem e suas aplicações na Neurociência:
Para desbloquear o próximo módulo, é necessário que você responda corretamente a uma das seguintes questões:
Clique aqui e retorne para saber como desbloquear.
Diferenciar os processos de aprendizagem motora e memória e aprendizagem associativa e não associativa
A aprendizagem motora pode ser concebida como o estabelecimento de um modelo interno que represente a correspondência exata entre as informações sensoriais e motoras percebidas.
Durante a fase inicial da aprendizagem motora, os movimentos não são qualificados, mas altamente dependentes de feedback e requerem grandes demandas de atenção.
Com prática, precisão e velocidade, as ações aumentam, enquanto o processamento de feedback se torna menos importante.
Em relação à neuroanatomia funcional, a aquisição de habilidades é acompanhada por mudanças em nível regional, assim como de circuito neural. Nos últimos anos, o desenvolvimento de técnicas de neuroimagem funcional forneceu informações sobre os mecanismos neuronais subjacentes às mudanças de comportamento durante a aquisição de habilidade motora.
É importante ressaltar que os dados de neuroimagem funcional fornecem uma melhor compreensão das observações clínicas. Vários fatores podem ser causa de discrepâncias nos resultados como, por exemplo, variações nas tarefas utilizadas, diferentes metodologias e a fase de aprendizagem motora.
Em geral, duas formas de aprendizagem motora podem ser distinguidas: aprendizagem explícita e implícita.
Aprendizagem explícita envolve a lembrança consciente de experiências anteriores.
Já a aprendizagem implícita é conhecida como uma forma não consciente de aprendizagem, caracterizada pelo aperfeiçoamento motor.
A habilidade motora melhora por meio do controle explícito que ocorre nos estágios iniciais de aprendizagem, alcançando um controle mais implícito ou automatizado nos estágios finais da aprendizagem do movimento.
Dessa forma, a aprendizagem motora é composta de três fases distintas:
Desempenho lento, movimentos irregulares, com tempo de atuação variável.
Aprendizagem gradual, com aumento da velocidade.
Desempenho rápido, automatizado e habilidoso, com movimentos finos.
Durante a fase inicial de aprendizagem por tentativa e erro, os sujeitos têm que descobrir o movimento correto.
O requisito crítico dessa fase é o novo estabelecimento de percepção de pistas sensoriais com os comandos motores corretos. Para isso, os sujeitos devem atender às pistas sensoriais, que decidem qual movimento inicializar a seguir, recebendo o feedback a partir da codificação do que é percebido em resposta.
Assim, o estabelecimento de uma associação sensório-motora arbitrária se faz necessária durante o aprendizado por tentativa e erro, que está intimamente relacionado a atenção, decisão e seleção de movimentos, processamento de feedback sensorial e memória de trabalho. Dessa maneira, quando os sujeitos descobrirem os movimentos corretos, o mapa de tradução sensório-motora é fixado.
Estímulos sensoriais devem ser retidos na memória de trabalho para serem traduzidos para a saída do motor – as ações desempenhadas ainda são lentas e não qualificadas, e o feedback e a atenção no processamento têm um papel crítico.
Com a prática sensório-motora, os mapas se tornam mais fortes e são armazenados na memória de longo prazo.
As dicas visuais são transformadas com precisão e rapidez para a necessidade de resposta motora. Portanto, a ação pode ser realizada com processamento de feedback sensorial menos intensivo e em alta velocidade. Após a prática de longo prazo, movimentos se tornam automáticos e podem ser executados em alta velocidade e precisão, mesmo que os sujeitos não prestem atenção à ação.
Ao olharmos para os mecanismos neurais subjacentes ao motor aprendizagem, surgem duas questões principais (WOOLLACOTT; SHUMWAY-COOK, 2010):
Pode-se perguntar sobre a contribuição de cada região do cérebro no processo de aprendizagem motora.
É interessante interpretar padrões de ativação que estão associados a estágios avançados de aprendizagem em termos de circuitos neurais.
Aprendizagem e memória são processos correlatos capazes de sofrer modificações com base em estímulos ambientais. Do ponto de vista processual, não é possível separar o aprendizado da memória, nem é possível essa distinção dentro do circuito neural. Assim, o que foi aprendido estaria relacionado à memória, sendo a expressão da capacidade de recuperar informações adquiridas. Portanto, os dois processos têm uma relação íntima.
No sistema nervoso, aprender significa produzir uma referência básica por meio de padrões neurais construídos e incorporados por vários estágios entre o nascimento e a maturidade.
A aprendizagem e a memória são incorporadas ao longo da vida, o que nos permite nos preparar para uma situação de resposta a um estímulo vivenciado anteriormente.
Nesse caso...
A aprendizagem é o processo pelo qual humanos e animais adquirem conhecimentos sobre o ambiente que os rodeia.
Já a memória seria a capacidade do indivíduo em reter e usar informações de maneiras diferentes e em períodos diferentes.
No início do século XX, Karl Lashley (1890-1958), psicólogo experimental, elaborou um dos primeiros modelos de memória distribuída. Esse modelo contrariava a visão da época, baseando-se na crença sobre a existência de um lugar específico no córtex responsável pelo armazenamento de informações. Essa posição foi defendida por pesquisadores renomados e que ficaram conhecidos como os “localizacionistas”.
Esse modelo teórico, em certa medida, ainda é utilizado, muito embora não tenha a intenção de encontrar um lugar onde estaria localizada a memória. Sendo assim, o que os cientistas procuram são as modificações na transmissão neuronal. No entanto, o objetivo de Lashley era, em princípio, encontrar um lugar da memória, mas ele alterou essa concepção ao longo de suas investigações.
Especificamente, Lashley desenvolveu um experimento que consiste em um labirinto onde dois grupos de ratos tiveram que encontrar a saída. Em um grupo, todos os ratos eram saudáveis e, apesar de algumas dificuldades iniciais, eles encontraram a saída. Com a repetição da tarefa, o grupo saudável aumentou a velocidade de execução, conseguindo sair do labirinto cada vez mais rápido, ao contrário do grupo com lesões cerebrais em diferentes partes do cérebro, que não evoluiu em nada. Lashley verificou que quanto maior a quantidade do tecido afetado, mais tempo demorava o animal para sair do labirinto, independentemente da localização da lesão.
Tendo em vista esse resultado, Lashley chegou à conclusão de que a memória estaria localizada por todo o sistema nervoso. Esses achados formaram a base da corrente antilocalizacionista da memória, propondo que diferentes áreas corticais contribuem igualmente para o processo de formação da memória.
Atualmente, já se sabe que Lashley interpretou mal seu experimento, uma vez que nem todas as áreas corticais contribuem da mesma forma para a memória. No entanto, suas descobertas foram extremamente importantes para a compreensão dos processos envolvidos no armazenamento de informações.
Donald Hebb (1904-1985), cientista canadense e principal discípulo de Lashley, propôs que um grupo de neurônios fosse ativado simultaneamente no córtex cerebral para representar um evento ou um objeto. Além de imaginar que os neurônios tinham que ser interconectados – sendo que nessa época as sinapses ainda não haviam sido descobertas –, o modelo de Hebb situava esse objeto em uma memória de curto prazo. Dessa forma, sua evocação repetida reforçaria as conexões entre esse grupo de neurônios. Após a aprendizagem, a ativação de apenas uma parte do circuito levaria à ativação de sua representação plena.
Em busca da descrição dos mecanismos mentais usados na memória, vários pesquisadores tentaram estabelecer uma sequência de processos.
Todo o processo mnemônico começa com a aquisição, que consiste na entrada de um evento no circuito neural ligado à memória. Geralmente, esses eventos são complexos, pois os sistemas de memória usam um processo de seleção e permitem apenas a aquisição de aspectos relevantes.
Apesar de esse processo de seleção ainda ser desconhecido, podemos exemplificá-lo facilmente ao tentar lembrar de uma jogada no contexto de uma partida de futebol. Nesse caso, só podemos lembrar de alguns momentos e não de toda a jogada. Após a aquisição, esses eventos são armazenados por algum tempo, que pode variar de alguns segundos a muitos anos. Esse fenômeno é chamado de retenção.
Em termos de tempo de retenção, podemos classificar a memória como memória de curta e longa duração:
Memória de curta duração tem uma capacidade de retenção de segundos, minutos ou horas e se refere ao momento presente, garantindo sua sequência lógica.
Já a memória de longo prazo é aquela que tem uma capacidade de retenção de horas ou dias, sendo responsável pela nossa história.
Como a memória, a aprendizagem não é um processo único e tem diferentes formas. Uma delas, considerada implícita, está associada com processos motores ou aprendizagem verbal, sem dependência de processos cognitivos.
As formas de aprendizagem implícita podem ser associativas ou não associativas.
A aprendizagem não associativa é caracterizada por resposta a um estímulo que ocorre com base no tempo e é observado por meio do comportamento. Existem dois tipos de aprendizagem não associativa:
Entendida como redução da eficiência do estímulo que, por não ser significativo, leva a ignorá-lo.
Intensificação da resposta com base em um forte estímulo capaz de produzir até a percepção de estímulos que, antes do evento, são considerados fracos.
Do ponto de vista neurofisiológico, as formas simples de aprendizagem implícita produzem mudanças na eficácia de transmissão sináptica.
Em experimentos realizados com moluscos, é possível observar mecanismos sensoriais associados à memória. Em particular, a aplicação de estímulos com borrifadas de água repetidamente no molusco produziu uma quebra nos potenciais sinápticos disparados pelo neurônio sensorial em interneurônios e neurônios motores (habituação). Esse fenômeno foi considerado como sendo o resultado da quebra do nível de glutamato liberado pela terminação pré-sináptica e capacidade reduzida das vesículas pré-sinápticas.
Essas vesículas contêm transmissores que impedem tanto seu movimento na zona ativa quanto sua disponibilidade para uma nova liberação. Essa redução na eficiência sináptica pode durar vários minutos, e podem ocorrer mudanças duradouras nas conexões sinápticas da área ativada (mudanças plásticas), produzindo armazenamento de uma memória reflexiva simples, que não é restrita a um único lugar, mas distribuída em um amplo circuito neural.
Nesse estudo, também foi possível observar que a aplicação de apenas um estímulo prejudicial para o molusco causou uma mudança nas conexões sinápticas do circuito neuronal de remoção reflexa das guelras, neurônios sensoriais, interneurônios ou neurônios motores. Esse problema deve ser levado em consideração, já que um único grupo de conexões sinápticas pode responder à habituação ou sensibilização.
Enquanto a habituação causa uma redução na força sináptica
A sensibilização leva a uma facilitação dessa força sináptica
Em outras palavras, facilitar o funcionamento dos neurônios melhora a liberação de transmissores nas sinapses de terminações de neurônios sensoriais.
Já a aprendizagem associativa é caracterizada por associações entre atos. Sendo assim, podemos diferenciar entre dois tipos de aprendizagem associativa:
Nele, a relação estímulo-resposta ocorre quando o primeiro estímulo evoca uma resposta que pode ser medida de forma diferente, sendo produzido como resultado da aplicação do segundo estímulo. O primeiro é o estímulo não condicionado, que não requer treinamento, nem preparação prévia para respostas, e atua com base na estimulação condicionada. Esse aprendizado não tem intervalo de tempo entre o estímulo condicionado e o não condicionado, fator fundamental para que ocorra uma convergência de estímulos nos neurônios sensoriais individuais. Essa peculiaridade promove uma facilitação pré-sináptica e sua intensidade de ações é inversamente proporcional ao intervalo de tempo aplicado. Essa facilitação é conhecida como potenciação de longo prazo (LTP).
Nele ocorre a associação de determinado comportamento a um estímulo significativo, uma relação estímulo-comportamento orgânica na qual o ato de dirigir é o resultado de uma resposta neurofisiológica para o sucesso. Baseia-se na tentativa e erro, ou seja, aprendemos a associar certa resposta, entre muitas que fizemos, com uma consequência.
O especialista Sergio Eduardo de Carvalho Machado apresenta as relações entre a aprendizagem e o processo de memorização:
Para desbloquear o próximo módulo, é necessário que você responda corretamente a uma das seguintes questões:
Clique aqui e retorne para saber como desbloquear.
Descrever os mecanismos cerebrais relacionados à recuperação de lesões neurológicas
A interpretação corriqueira de sinais anormais após uma lesão do sistema nervoso central (SNC) é que o desfecho está diretamente associado aos danos no SNC:
Uma lesão em qualquer local específico dá ascensão a um conjunto definido de sinais físicos.
Inversamente, uma constelação particular de sinais físicos indica uma lesão em um local específico do SNC.
Está implícito nessa interpretação que o resto do SNC continua funcionando, no entanto, com alguns déficit. Se o SNC fosse composto de um tipo simples de fiação de central telefônica, então essa seria uma visão razoável. Porém, esse não é o caso.
Por exemplo, se fizermos uma lesão no cerebelo de um macaco, a lesão resultará em tremor e alteração no tempo dos movimentos. Com a recuperação, o posicionamento dos membros (dependente no córtex somatossensorial) torna-se de maior importância e o tremor pode ser mascarado. Se uma lesão agora for produzida no córtex somatossensorial, o deficit cerebelar original reaparecerá e se tornará permanente.
Parece que pelo menos parte do quadro clínico de uma lesão não é devido à lesão em si, mas à maneira pela qual o SNC intacto (ou seja, não lesionado) altera como uma resposta à lesão, ou seja, em como o SNC se reorganiza após uma lesão. Em suma, sob certas condições, uma lesão no SNC pode induzir células nervosas intactas e seus axônios a fazerem sinapses com áreas que perderam suas aferências normais após a lesão.
Esse rearranjo de conexões é talvez um dos mecanismos mais importantes subjacentes à recuperação da função e tem grandes implicações, visto que, claramente, no SNC alterado, as vias que estavam relativamente sem uso ou pouco usadas agora se tornarão eficazes.
No entanto, essa conectividade alterada também pode ser responsável por deficit permanente e comprometimento da função. No local da lesão, uma sequência bem conhecida de eventos acontece. Embora se fale muito no local da lesão no SNC, a visão geral é que um pequeno número de axônios possa regenerar-se após a lesão. Alguma recuperação funcional pode ser devido à recuperação de células nervosas parcialmente danificadas e por remielinização.
As doenças neurológicas são uma das principais fontes de incapacidade e mortalidade no mundo. Algumas das mais significativas incluem acidente vascular encefálico (AVE), esclerose múltipla, traumatismo crânio encefálico e lesão medular. Em todos esses casos, e em muitos outros, o processo da doença pode causar diversos deficit sensório-motores, com uma melhora espontânea ocorrendo apenas meses depois. No entanto, essa melhora é muitas vezes insuficiente para restaurar a função satisfatoriamente e tem levado muitos pesquisadores e clínicos a buscar métodos pelos quais a melhoria pode ser acelerada e maximizada.
Em cada doença mencionada, os deficit são causados por disfunções neuronais. A recuperação pode, portanto, ocorrer por múltiplos mecanismos como:
A restauração da função dos neurônios danificados
A mudança na estrutura e função dos neurônios não danificados remanescentes, assim como das glias relacionadas
As células da glia são células não neuronais do sistema nervoso central que proporcionam suporte e nutrição aos neurônios.
A formação de novas estratégias comportamentais para cumprir as funções afetadas
A recuperação metabólica dos neurônios provavelmente ocorrerá ao longo de alguns dias, portanto, muito da recuperação que é vista após esse período de dias é provável que represente um dos dois últimos mecanismos.
Embora a capacidade de mudar o comportamento tenha sido reconhecida como tendo uma base neural, os mecanismos pelos quais tal plasticidade ocorre foram descobertos apenas nas últimas décadas. Isso começou com a descoberta da potencialização de longo prazo (LTP) no hipocampo – e depois em outras áreas, incluindo o córtex motor.
Mais recentemente, processos complementares foram descobertos, mostrando que eles mantêm a atividade neuronal em face da alteração da atividade sináptica.
Esses mecanismos homeostáticos têm o potencial de restaurar a função neuronal, mesmo quando uma grande parte da entrada sináptica é perdida. Esses mecanismos são susceptíveis de reverter o fenômeno de diásquise, ou perda da função neuronal devido à desaferentação parcial. No entanto, a contribuição da diásquise para os deficit sensório-motores ainda é pouco clara.
Um exemplo mais prático de mudanças ocorridas no mapa cortical ocorre em indivíduos amputados. Geralmente, as mudanças corporais nesses indivíduos fazem com que o membro fantasma mantenha uma relação natural com o coto.
Ação exercida pela lesão cerebral que causa uma perturbação difusa na região localizada ao seu redor.
Perda da inervação sensorial de uma região.
Parte do membro que permanece após a cirurgia de amputação.
Membros fantasmas indolores, especialmente do braço, sofrem telescopagem gradual em que o braço e antebraço gradualmente desaparecem da consciência, isto é, evolui com redução progressiva do tamanho do membro, ficando a sensação de que a mão e os dedos se aproximam do coto, ou até mesmo a sensação de penetrarem no coto. Dor de gravidade variável, de incômoda à altamente angustiante, pode envolver o fantasma, o coto ou ambos. Os membros fantasmas também ocorrem após a avulsão do plexo braquial, e após a transecção da medula espinhal com paraplegia.
O fenômeno da telescopagem acomete pacientes com sensação de membro fantasma. Nele, há alteração sinestésica do membro amputado – se antes ele era sentido em sua totalidade, com o tempo o tamanho do membro fantasma vai diminuindo, podendo chegar ao membro residual como, por exemplo, o coto.
Sobre o membro fantasma, é comum o relato de pacientes que continuam sentindo o membro mesmo após a amputação. Essa sensação é denominada “síndrome do membro fantasma” e acomete principalmente amputados de origem traumática, como acidentes de trânsito. Vale ressaltar que cerca de 70% dos amputados podem senti-la ao longo da vida.
Em relação ao mecanismo neural subjacente a um membro fantasma, ele é criado pela má adaptação que ocorre na atividade do córtex parietal, que atua na regulação da somatossensação na parte amputada. Pode-se esperar que um padrão neural doloroso de pré-amputação possa coexistir.
Outras evidências de reorganização sensorial cortical em amputados mostram que sensações de toque na face e no coto foram referidas somatotopicamente para regiões particulares do membro fantasma.
O aparecimento do fenômeno quatro semanas após a amputação foi considerada como evidência contra o brotamento como base da reorganização, e a favor do desmascaramento do silêncio preexistente das conexões sensoriais.
Em outros experimentos, o médico neurologista Vilayanur Ramachandran, usando uma caixa de espelho, proporcionou a indivíduos amputados a movimentação de seus dedos fantasmas imóveis por meio de imagem espelhada de sua própria mão normal em movimento, quando o membro fantasma foi sobreposto visualmente. Em outro experimento, Ramachandran, também usando a caixa de espelho, aplicou estímulos sensoriais à mão normal de um indivíduo, enquanto o mesmo observava pelo espelho, sentindo as sensações no mesmo lugar no membro fantasma.
Esses resultados sugerem que há uma quantidade considerável de plasticidade latente no cérebro adulto, com caminhos ligando os dois hemisférios, emergindo rapidamente (BEAR et al., 2008).
A maioria das pessoas que sobrevive a um acidente vascular, por exemplo, experimenta alguma recuperação motora, funções sensoriais e/ou cognitivas nos meses seguintes. Acredita-se que essa recuperação funcional é o efeito de alguma forma de reorganização do sistema nervoso central que ocorre após o dano cerebral.
Até algumas décadas atrás, pouco se sabia sobre esses fenômenos de reorganização cerebral ou neuroplasticidade. Com a introdução das técnicas de neuroimagem funcional, como a RMf e a TEP, foi possível alcançar novos conhecimentos sobre os mecanismos cerebrais oriundos da recuperação funcional.
Desde os primórdios da neurologia, o estudo da relação entre anatomia e função sempre esteve presente e gerou conflitos. Apesar do surgimento de diversas teorias, muitas delas errôneas e generalistas, atualmente sabe-se que o cérebro de fato apresenta relações com diversas áreas corporais distribuídas em diferentes pontos do cérebro, principalmente para as atividades motoras e sensitivas. Desse modo, o córtex tem uma representação detalhada e organizada de toda a superfície corporal, assim como do campo visual, no caso dos sistemas sensoriais.
O avanço das técnicas investigativas permite isolar áreas muito específicas e relacionar suas funções. A distribuição somatotópica no córtex sensitivo primário (S1) tem uma distribuição muito precisa, no entanto, no córtex motor primário (M1), a distribuição é mais complexa. Existem nele vias convergentes de diversas áreas cerebrais para um único músculo, dando o substrato para as diferentes combinações de estratégias de movimento.
Distribuição precisa
Distribuição complexa
Há também vias divergentes de uma única área para diversos músculos e muitas interconexões horizontais entre sub-regiões da M1, indicando uma complexa interação para a realização de movimentos multiarticulares ou multissegmentares, como a estabilização proximal de um braço para o sucesso de uma atividade independente de um dedo da mão.
Esses elementos são uma peça importante para a plasticidade neural. Diferentes áreas representativas para uma mesma função motora podem servir como uma via substituta de outra área lesionada, dando maior flexibilidade às readaptações.
Diversos estudos têm demonstrado que, após a lesão da área M1, áreas motoras secundárias, como o córtex pré-frontal dorsolateral (CPFDL), área motor suplementar (AMS) e parte posterior do giro cingulado, são utilizadas para a recuperação motora.
Pacientes com pouco deficit motor apresentam pouca remodelação cortical.
Enquanto outros, com alto grau de incapacidade, têm maiores porções de áreas secundárias ativadas.
O termo plasticidade cerebral abrange os possíveis mecanismos de reorganização neuronal como: o recrutamento de vias funcionalmente homólogas, mas anatomicamente distintas das áreas lesionadas, sinaptogênese, arborização dendrítica e ativação de áreas secundárias vistas no desmascaramento, ou seja, conexões que sob circunstâncias normais são pouco utilizadas, mas respondem fortemente após a lesão cerebral, refletindo uma redundância nas conexões corticais.
Processo de formação de sinapses entre os neurônios do sistema nervoso central.
Brotamento de novos terminais axônicos
Há ainda dúvidas quanto aos efeitos da lesão ao hemisfério cerebral contralateral na recuperação motora. Em geral, na fase aguda há uma diminuição da responsividade do hemisfério lesionado e um aumento da atividade no hemisfério contralateral à lesão – e esta atividade é maior nos pacientes com maior comprometimento motor.
Estudos realizados com pacientes que sofreram hemisferectomia demonstram que esses indivíduos experimentam algum grau de recuperação, sendo que a estimulação cortical do hemisfério remanescente induziu potenciais motores em ambos nos músculos deltoides e bíceps esquerdo e direito.
Fase da doença que surge após a infecção/lesão, na qual os sintomas clínicos são mais nítidos. É um período de definição: o paciente se cura, passa para a fase crônica ou morre.
Remoção de todo ou quase todo o hemisfério cerebral.
Como o tempo de resposta à estimulação cortical é mais longo, é possível que as ações do hemisfério não afetado sobre músculos ipsilaterais ocorram por meio das vias polissinápticas, como o trato reticuloespinhal.
Já em outros estudos a atividade do hemisfério não lesionado não foi relacionada com a melhora da função motora contralateral e ainda pode atuar de forma inibitória no hemisfério contralateral, por meio de modulação por neurônios GABAérgicos, reduzindo a excitabilidade cortical. Em pacientes com hemiparesia após AVE, essa forma de inibição recíproca pode ser um obstáculo para o hemisfério lesionado, sendo essa inibição mais evidente nos indivíduos com maior incapacidade motora.
Ainda não é possível determinar se a atividade no hemisfério não lesionado atua de forma compensatória após a lesão, ou se são apenas as vias secundárias que só se tornam aparentes após a lesão de um dos hemisférios.
Esses achados indicam que pacientes que demonstram recuperação relacionada ao hemisfério lesionado têm melhor prognóstico que aqueles em que a possível recuperação se dá pelo hemisfério não lesionado, e quanto maior for a simetria da distribuição funcional após lesão de um hemisfério, melhor o prognóstico.
A reorganização cerebral inicia-se ainda na fase aguda, após a lesão cortical, com melhora progressiva até a estabilização do quadro clínico. Apesar das remodelações corticais serem mais proeminentes nas fases iniciais, mudanças na atividade cerebral continuam até um ano após a lesão.
Estudos longitudinais que avaliaram alterações nas áreas motoras compensatórias de pacientes com hemiplegia discordam quanto aos resultados:
(Hemi - metade, plegia - paralisia)
É a paralisia de metade sagital (esquerda ou direita) do corpo.
Alguns demonstrando um aumento inicial da área recrutada seguida de uma redução progressiva dessas áreas, correlacionadas com a recuperação motora.
Enquanto outros demonstraram um aumento progressivo da área secundária ativada.
O processo de aprendizado motor é extremamente dependente da frequência de experimentação ou prática de uma tarefa. Em indivíduos normais, alterações corticais transitórias podem ser vistas em poucas horas de treino repetitivo, como determinado movimento de um dedo, por exemplo, resultando no aumento de sua área cortical representativa.
Alguns estudos demonstram um aumento da área cerebral de representação motora imediatamente após uma sessão de fisioterapia em pacientes que foram acometidos por AVE e, no dia seguinte à intervenção, a área diminuiu novamente.
A reorganização de longo prazo necessita de um maior período de treinamento, por exemplo, quatro semanas, com suas mudanças persistindo por vários meses. Em pacientes com AVE subcortical, o treino intensivo durante três semanas aumenta a ativação dos córtices M1 e S1 no hemisfério lesionado, comparado com os tratamentos padrão.
Essas mudanças são principalmente mediadas por glutamato, sendo que o aumento da excitabilidade do hemisfério contralateral e de regiões ao redor de uma área que sofreu isquemia pode ser resultado da diminuição da atividade inibitória GABAérgica e de alterações neuroprotetoras relacionadas ao pré-condicionamento isquêmico, que pode aumentar a susceptibilidade de mudanças adaptativas relacionadas à recuperação de funções corticais vista em estudos de RMf e TEP.
Com relação à recuperação, o feedback proprioceptivo é extremamente positivo durante o processo. A realização de movimentos passivos induz, em indivíduos normais, uma ativação sensório-motora semelhante ao movimento voluntário, com exceção de áreas no giro do cíngulo e nos núcleos da base, e induzem uma reorganização cortical após algumas semanas de realização.
É a falta de fornecimento sanguíneo para um tecido orgânico devido à obstrução causada por um trombo, seja ele formado por placas gordurosas ou por coágulos sanguíneos.
A resposta motora também poder ser facilitada pelo treino mental de movimentos não acompanhados de movimento, a chamada prática mental (PM).
A PM consiste em um método de treinamento pelo qual a reprodução interna de determinado ato motor (simulação mental) é repetida extensivamente com a intenção de promover a aprendizagem ou melhoria das habilidades motoras. Essa simulação mental (imagens motoras) corresponde a um estado dinâmico durante a execução de uma ação específica, reativado internamente na memória de trabalho na ausência de qualquer movimento.
A PM resulta de um acesso consciente à intenção motora, que geralmente é realizada inconscientemente durante a preparação motora, estabelecendo uma relação entre eventos motores e percepções cognitivas. Tal técnica é muito utilizada no treinamento esportivo de alta performance, mas também pode ser utilizado nos pacientes com deficit na movimentação.
Estudos têm mostrado que há semelhança nas funções psicofísicas e fisiológicas entre movimentos reais e imaginários, sugerindo que haja processos comuns entre ambos. Experimentos usando RMf mostraram que não apenas as áreas motoras suplementares, pré-motoras e cerebelo foram ativadas durante movimentos imaginários da mão e dos dedos mas, também, a M1 contralateral.
Em relação à semelhança das áreas corticais ativadas e engajadas na execução e imaginação de um movimento, foi descoberto que tais semelhanças na atividade cerebral ocorreram durante os movimentos dos dedos.
Dentro desse contexto, várias investigações tentam aplicar esses achados no processo de reabilitação motora, especialmente nos casos que dizem respeito à recuperação de áreas envolvidas na função motora, após AVE.
A PM aplicada sozinha alcança resultados menos interessantes do que a reabilitação motora isolada em pacientes que foram acometidos por AVE. No entanto, a combinação de ambas as técnicas (PM e reabilitação motora) mostrou-se mais eficaz do que as duas técnicas aplicadas sozinhas.
A prática mental representa uma abordagem interessante para a reabilitação motora de pacientes pós-AVE em todos os estágios da recuperação:
Ao contrário das terapias motoras ativas e passivas, a PM, em princípio, não depende da função residual.
Em pacientes pós-AVE, a PM pode ser usada como um substituto para o movimento executado como uma forma de ativar as redes neurais responsáveis pelas funções motoras. Assim, a PM é mais eficaz quando as habilidades que estão sendo ensaiadas mentalmente também são praticadas fisicamente. Isso se deve a um "esquema motor" que a PM reforçaria ao longo do treinamento.
O especialista Sergio Eduardo de Carvalho Machado apresenta as evidências sobre o papel do exercício na Neuroplasticidade:
Para desbloquear o próximo módulo, é necessário que você responda corretamente a uma das seguintes questões:
Clique aqui e retorne para saber como desbloquear.
Com relação aos campos da Neurociência, conforme apontado, estão envolvidas diversas áreas de atuação, como Matemática, Física, Linguística, Engenharia, Ciência da Computação, Química, Filosofia, Biologia, Psicologia, Educação Física, Medicina, mostrando o quão complexa e multidisciplinar é a área.
Conhecemos as principais técnicas de neuroimagem, como a RMf e TEP, e o quanto são importantes para investigar a conectividade cortical em função de diferentes tipos de tratamento, como medicamentos, técnicas de reabilitação física e cognitiva, exercício físico, compreendendo as variáveis neurofisiológicas relacionadas com a aplicação dessas técnicas e proporcionando melhor compreensão quanto ao funcionamento e reorganização do cérebro que ocorre por mecanismos como o "surgimento axonal", no qual axônios não danificados desenvolvem novas terminações nervosas para reconectar os neurônios cujas ligações foram danificadas ou rompidas.
Os axônios não danificados também podem brotar terminações nervosas e se conectar com outras células nervosas não danificadas, formando novas vias neurais para realizar uma função necessária.
Vimos ainda os mecanismos e processos relacionados à aprendizagem motora e memória, e aprendizagem associativa e não associativa, importantíssimos para entender como ocorre a recuperação de lesões.
Agora, o especialista Sergio Eduardo de Carvalho Machado encerra este conteúdo com as abordagens mais atuais de estudos na área da neurociência. Ele fala sobre o paradigma de que no cérebro não existe neurogênese após seu amadurecimento e o papel do hipocampo nesse processo; expõe a recuperação de lesões com sequelas motoras, abordando os estudos na área de Biofeedback e eletroestimulação para a plasticidade neuronal. Finaliza esclarecendo como o processo de novas aprendizagens (habilidades) podem ajudar a neuroplasticidade.
 / Wikimedia Commons / Domínio Público.")



.")


.")
.")
