Descrição
Principais funções e morfologia básica dos tecidos cartilaginoso, ósseo e muscular.
PROPÓSITO
Compreender as características morfológicas e funções do tecido muscular e de duas outras especializações do tecido conjuntivo, os tecidos cartilaginoso e ósseo, é importante para iniciar os estudos correlacionados à histologia aplicada, aos mecanismos fisiológicos dos órgãos do corpo e para entender as bases patológicas das doenças.
OBJETIVOS
Módulo 1
Descrever as características morfológicas, a função e a localização do tecido cartilaginoso, bem como os principais tipos de cartilagem e as suas particularidades
Módulo 2
Descrever as características morfológicas do tecido ósseo, os componentes fundamentais, a estrutura geral e a formação dos diferentes tipos de ossos
Módulo 3
Reconhecer os três tipos de tecido muscular, as suas características morfológicas e as diferentes formas de contração muscular
Introdução
No corpo humano, existem cerca de 100 trilhões de células, que apresentam as mais variadas morfologias e funções. Essas células se aglomeram em vários grupos específicos e constituem os quatro principais tecidos do corpo (epitelial, conjuntivo, muscular e nervoso), que formam os órgãos e garantem a manutenção do organismo.
Neste conteúdo, estudaremos o tecido cartilaginoso, que é uma das especializações do tecido conjuntivo. Veremos os tipos de cartilagem, suas características e localização, além dos tipos de crescimento cartilaginoso e reparo tecidual. Em seguida, ainda sobre as especializações do tecido conjuntivo, estudaremos o também importante tecido ósseo e os seus componentes. Aprenderemos os tipos de ossos, a sua estrutura básica, o processo de ossificação e, por último, as cartilagens. Por fim, conheceremos o tecido muscular, um dos quatro tecidos básicos do corpo. Veremos com detalhes o músculo estriado esquelético, o estriado cardíaco e o músculo liso, bem como os mecanismos moleculares envolvidos no processo de contração celular.
AVISO
Em nosso material, unidades de medida e números são escritos juntos (ex.: 25km) por questões de tecnologia e didáticas. No entanto, o Inmetro estabelece que deve existir um espaço entre o número e a unidade (ex.: 25 km). Logo, os relatórios técnicos e demais materiais escritos por você devem seguir o padrão internacional de separação dos números e das unidades.
MÓDULO 1
Descrever as características morfológicas, a função e a localização do tecido cartilaginoso, bem como os principais tipos de cartilagem e as suas particularidades
Características gerais e funções do tecido cartilaginoso
Como vimos na introdução, o tecido cartilaginoso é uma das especializações do tecido conjuntivo e, como tal, é formado por células e matriz extracelular. Esses componentes são envolvidos pelo pericôndrio, que é formado por uma camada de células indiferenciadas que podem vir a se diferenciar em condroblastos. O tecido cartilaginoso fornece suporte aos tecidos moles, reveste as superfícies articulares, absorvendo o choque, e facilita o deslizamento dos ossos nas articulações.
Assim como os fibroblastos e adipócitos (nos tecidos conjuntivo e adiposo, respectivamente), os condroblastos derivam das células mesenquimais. Eles apresentam retículo endoplasmático bem desenvolvido e reservas de lipídeos e glicogênio. Quando maduros, os condroblastos são denominados de condrócitos.

A proliferação dos condroblastos resulta no crescimento da cartilagem. Na cartilagem adulta, os condrócitos raramente se dividem, mas sintetizam continuamente as moléculas necessárias para a renovação constante da matriz extracelular, especialmente os proteoglicanos. Por conta disso, os condrócitos possuem necessidades nutricionais significativas. Os condrócitos ocupam pequenas cavidades na matriz extracelular chamadas de lacunas. Podemos encontrar dois condrócitos ocupando uma única lacuna. Na figura a seguir, vemos um corte histológico do tecido cartilaginoso. Observe as lacunas encontradas nele.

A matriz do tecido cartilaginoso é constituída basicamente por colágeno ou colágeno e elastina, em associação com macromoléculas de proteoglicanos, ácido hialurônico e diversas glicoproteínas. Além disso, é compartimentalizada: uma matriz pericelular que é rodeada por uma matriz territorial. A camada mais interna da matriz territorial é a borda lacunar, formada por fibrilas colágenas frouxas. A matriz territorial é formada por fibrilas de colágeno do tipo II, aleatoriamente dispostas e circundadas por proteoglicanos, como mostrado na figura ao lado.


A rede de fibrilas de colágeno é resistente à tensão, e os agregados de proteoglicanos, altamente hidratados, são muito sensíveis às forças de cisalhamento. A forte interação entre essas duas classes de moléculas, com características contrastantes, torna a cartilagem bem adaptada para sustentação de peso, como podemos observar na figura ao lado, principalmente em pontos de movimento, como as articulações sinoviais. Como essa propriedade é mantida durante o crescimento do organismo, a cartilagem é essencial para o desenvolvimento, desde o esqueleto fetal até a maioria dos ossos em crescimento.
A consistência firme das cartilagens é consequência das ligações eletrostáticas entre os glicosaminoglicanos e o colágeno, e da grande quantidade de moléculas de água presas a esses glicosaminoglicanos, principalmente.
Em contraste com o tecido conjuntivo propriamente dito, o tecido cartilaginoso não contém vasos sanguíneos, e sua nutrição ocorre a partir dos capilares presentes no pericôndrio. As cartilagens que revestem as superfícies dos ossos nas articulações móveis não possuem pericôndrio, e o aporte de nutrientes é dado pelo líquido sinovial das cavidades articulares. Além disso, é importante comentarmos que o tecido cartilaginoso também não apresenta vasos linfáticos e nervos.
Tipos de cartilagem
A cartilagem hialina é o tipo de cartilagem mais frequentemente encontrado no corpo humano. Ela forma o primeiro esqueleto do feto, que é gradativamente substituído por um esqueleto ósseo a partir de um processo conhecido como ossificação endocondral, que veremos mais adiante.
Então, ao longo do processo de desenvolvimento, a maior parte do modelo cartilaginoso é substituída por osso.
Apesar disso, o tecido cartilaginoso ainda persiste nas extremidades proximais e distais do osso, atuando como locais de crescimento dele. Essas regiões, chamadas de discos epifisários, apresentam cartilagem funcional enquanto o osso estiver crescendo. Cessado o crescimento em um indivíduo adulto, a cartilagem fica restrita à superfície das articulações móveis (cartilagem articular) e ao esqueleto da caixa torácica (cartilagens intercostais). A cartilagem hialina também é encontrada na parede das fossas nasais, na traqueia, na laringe e nos brônquios.
A cartilagem hialina é formada majoritariamente por delicadas fibrilas de colágeno do tipo II, associadas a moléculas de ácido hialurônico, proteoglicanos hidratados e glicoproteínas. Dentre as glicoproteínas estruturais, a condroitina é muito importante, pois apresenta sítios de ligação para condrócitos, fibras colágenas e glicosaminoglicanos, auxiliando na formação do arcabouço de matriz com os condrócitos. Na figura a seguir, observamos como essas estruturas estão organizadas.

Ao redor dos condrócitos existem regiões estreitas, ricas em proteoglicanos e pobres em colágeno, chamadas indevidamente de cápsulas. A matriz da cartilagem hialina é basofílica, ou seja, cora-se em azul com a hematoxilina durante as preparações histológicas, devido aos glicosaminoglicanos sulfatados (apresentam carga negativa).
Com exceção das articulares, todas as cartilagens hialinas são envolvidas pelo pericôndrio, que é uma camada de tecido conjuntivo. Como você deve lembrar, o tecido cartilaginoso é desprovido de capilares sanguíneos e, por isso, o pericôndrio é responsável pela nutrição, oxigenação e eliminação dos resíduos do metabolismo da cartilagem. Os nutrientes presentes no sangue atravessam o pericôndrio, penetram na matriz cartilaginosa e alcançam os condrócitos mais profundos. Por conta da característica avascular, a espessura máxima das cartilagens é limitada.
O pericôndrio é rico em fibras de colágeno do tipo I na parte mais superficial, apresentando mais células à medida que se aproxima da cartilagem. As células do pericôndrio mais próximas da cartilagem são capazes de se multiplicar e dar origem aos condroblastos e, posteriormente, aos condrócitos. Observe na figura a seguir como os condrócitos (às vezes dois) ocupam as pequenas cavidades da abundante matriz extracelular.
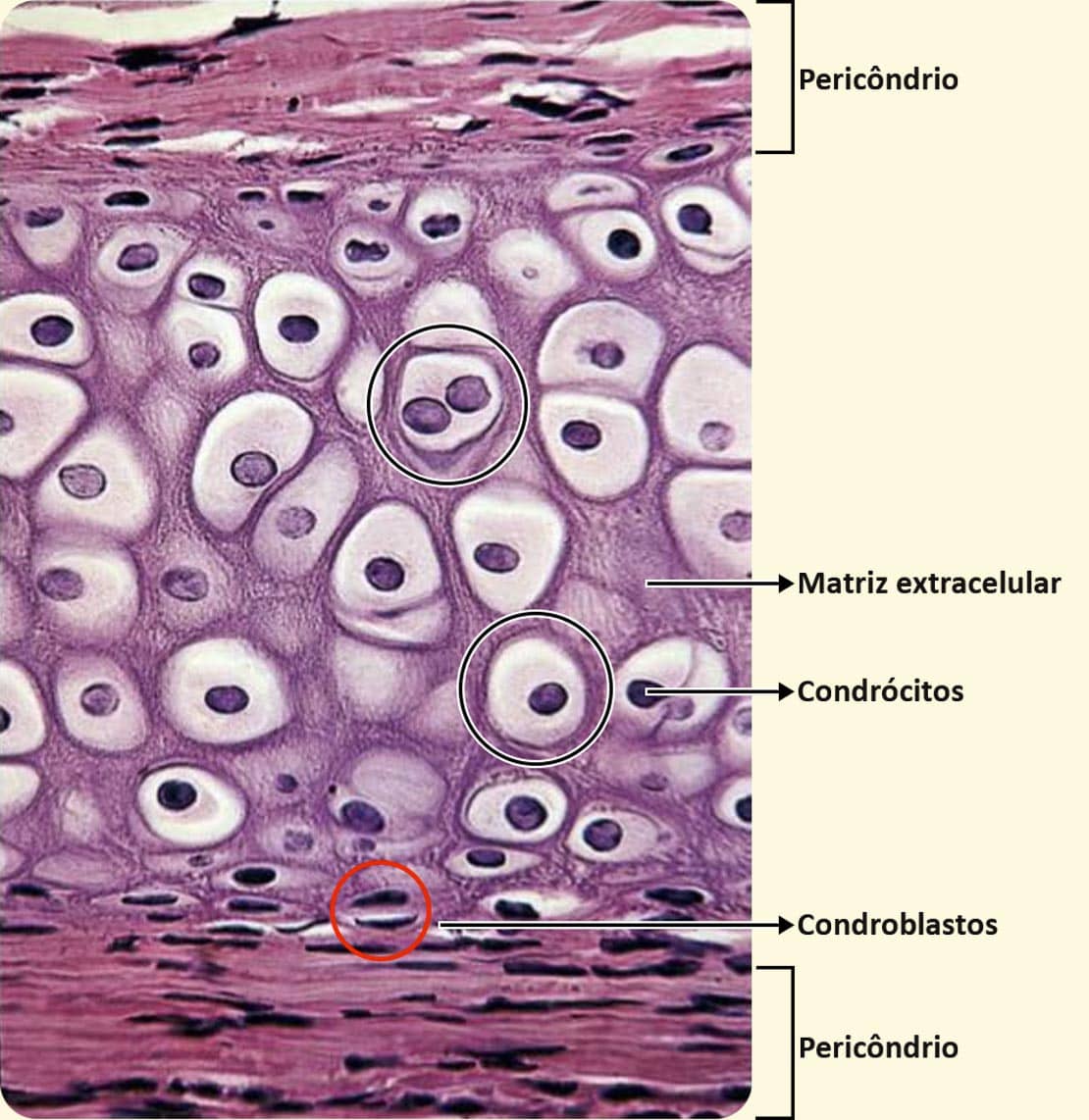
Os condrócitos secretam colágeno (principalmente do tipo II), proteoglicanos e glicoproteínas, como a condronectina. Apresentam morfologia alongada na periferia da cartilagem e arredondada nas regiões mais profundas. De um único condroblasto derivam grupos de até oito células, os chamados grupos isógenos, como mostrado na figura ao lado.
Embora pareça regular ao microscópio de luz, a superfície dos condrócitos apresenta reentrâncias e saliências, mais frequentes nas células mais jovens. Essa característica aumenta a superfície de contato celular, facilitando as trocas de nutrientes e metabólitos com o meio extracelular.

Você sabia
Tanto as células quanto a matriz cartilaginosa sofrem retração durante as técnicas de processamento histológico e, por isso, os condrócitos aparecem com uma forma estrelada e afastados da cápsula ao microscópio. Já em tecidos vivos e cortes bem preparados, essas células ocupam totalmente as lacunas da matriz.
Cartilagem elástica
A cartilagem elástica, observada na figura a seguir, é semelhante à hialina, porém, além das fibras de colágeno (especialmente do tipo II) interagindo com proteoglicanos, apresenta elastina em sua matriz, formando uma abundante rede de fibras elásticas ramificadas contínuas com as do pericôndrio. Essa característica oferece elasticidade, resiliência e maleabilidade à cartilagem.
Resiliência
Propriedade que alguns corpos apresentam de retornar à forma original após terem sido submetidos a uma deformação elástica.
Podemos encontrar a cartilagem elástica na tuba auditiva, na epiglote, no pavilhão auditivo, na orelha externa e na cartilagem cuneiforme da laringe. Diferentemente da cartilagem hialina, a matriz da cartilagem elástica não se calcifica durante o envelhecimento.

Fibrocartilagem
Também chamada de cartilagem fibrosa, a fibrocartilagem é considerada um tecido intermediário entre a cartilagem hialina e o tecido conjuntivo denso, estando sempre associada a esse último.
Possui grande força tensora, formando grande parte dos discos intervertebrais, sínfise pubiana, discos articulares do joelho, mandíbula e locais de inserção de tendões e ligamentos a ossos. Observando detalhadamente esses locais, notamos que a fibrocartilagem atua como um amortecedor de choques, conferindo aos tecidos resistência à compressão e às forças de cisalhamento.
As células da cartilagem fibrosa sintetizam várias moléculas da matriz, tanto em seu estado de desenvolvimento quanto em seu estado maduro totalmente diferenciado, como observado na figura a seguir. Dessa forma, a fibrocartilagem está preparada para as mudanças no ambiente externo, como forças mecânicas, alterações nutricionais, de hormônios e fatores de crescimento.

A matriz da fibrocartilagem é bem acidófila (com a eosina utilizada nas preparações histológicas, cora-se em rosa) por conter grandes quantidades de fibras colágenas. As fibras são principalmente de colágeno do tipo I, mas também são encontradas fibras de colágeno do tipo II que circundam os condrócitos e fibroblastos. As numerosas fibras formam feixes com orientação aparentemente irregular entre as células ou arranjos paralelos ao longo dos condrócitos em fileiras, como observado na figura ao lado. Não existe pericôndrio.

Você sabia
A razão entre os tipos de colágeno na fibrocartilagem se modifica com a idade. Em indivíduos idosos, há uma maior quantidade de colágeno do tipo II por conta da atividade metabólica dos condrócitos, que produzem e liberam constantemente essas fibrilas colágenas na matriz circundante.
A substância fundamental é bem reduzida, estando limitada às proximidades das lacunas que contêm os condrócitos. Apresenta quantidades maiores de versicano (proteoglicano secretado por fibroblastos) que agrecano (secretado por condrócitos). O versicano liga-se ao ácido hialurônico, formando agregados altamente hidratados. A degeneração dos discos intervertebrais está associada à degradação proteolítica desses agregados, que ficam na matriz da cartilagem.
Substância fundamental
Complexo de ácido hialurônico, proteoglicanos e glicoproteínas.
Agora que já comentamos as características gerais da fibrocartilagem, aprofundaremos um pouco mais nosso estudo abordando os discos intervertebrais.
Discos intervertebrais
Estão localizados entre os corpos das vértebras, unidos a elas por ligamentos. São formados por dois componentes:
É constituído por tecido conjuntivo denso e fibrocartilagem em maior quantidade, cujos feixes de colágeno formam camadas concêntricas.
É formado por células arredondadas, dispersas em um líquido viscoso rico em ácido hialurônico e com pouca quantidade de colágeno tipo II.
Na figura a seguir, conseguimos observar um esquema dos discos intervertebrais. Note que o núcleo pulposo está na parte central circundado pelo anel fibroso.

Por ser altamente hidratado e lubrificado, os discos intervertebrais funcionam como uma espécie de almofada, protegendo as vértebras contra impactos e prevenindo o desgaste vertebral durante os movimentos da coluna espinal.
Saiba mais
Em indivíduos jovens, o núcleo pulposo é mais abundante, sendo parcialmente substituído por fibrocartilagem com a idade.
Crescimento e reparo da cartilagem
O processo de desenvolvimento da cartilagem, a condrogênese, se dá no mesênquima, onde as células mesenquimais ganham morfologia arredondada, retraem seus prolongamentos e se multiplicam, formando aglomerados celulares chamados de nódulo condrogênico.
Com a expressão do fator de crescimento do tipo SOX-9, inicia-se o processo de diferenciação dessas células em condroblastos, que começam a secretar matriz cartilaginosa. À medida que a matriz vai sendo secretada e depositada, os condroblastos vão se afastando uns dos outros. A diferenciação das cartilagens ocorre do centro para a periferia: enquanto as células periféricas ainda são condroblastos típicos, as células mais centrais já apresentam as características de condrócitos. O mesênquima que circunda o nódulo condrogênico dá origem ao pericôndrio.
Fator de crescimento
Substância normalmente de origem proteica que funciona como importante molécula e comunicação entre as células, importante para a diferenciação celular.
A figura a seguir apresenta como ocorre o crescimento da cartilagem hialina.

Como vista na figura anterior, a partir do mesênquima (A) a multiplicação das células mesenquimais constitui um tecido formado por muitas células (B). Em seguida, com a secreção da matriz, os condroblastos se afastam (C). Por fim, a multiplicação mitótica dessas células dá origem aos grupos isógenos (D).
A regulação da condrogênese se dá pela ação de muitas moléculas, como receptores nucleares, fatores de transcrição, moléculas de adesão e proteínas da matriz. Forças biomecânicas também atuam no desenvolvimento do esqueleto cartilaginoso, não apenas regulando o formato, a regeneração e o envelhecimento da cartilagem, como também modificando as interações das células com a matriz cartilaginosa.
Com o início da secreção da matriz, o crescimento da cartilagem continua a partir de dois processos:
É um processo no qual a nova cartilagem é formada na superfície de uma cartilagem existente, como observado na figura a seguir. As células mais internas do pericôndrio, semelhantes a fibroblastos, produzem o colágeno do pericôndrio (do tipo I) e, uma vez que o crescimento cartilaginoso é iniciado, sofrem um processo de diferenciação para condroblastos, orientado pelo fator de transcrição SOX-9, que já comentamos. A nova matriz vai aumentando a massa cartilaginosa ao mesmo tempo que novos fibroblastos vão sendo produzidos para manter a população do pericôndrio.

Denomina o processo de formação da nova cartilagem dentro de uma massa de cartilagem existente. As novas células originam-se das mitoses de condrócitos dentro das lacunas da matriz, que são distensíveis e permitem a atividade secretora adicional. Inicialmente, as células filhas ocupam a mesma lacuna, mas com a secreção da nova matriz, cada uma passa a ocupar sua própria lacuna, afastando-se ainda mais com a contínua secreção da matriz.
O crescimento intersticial apresenta ocorrência quase exclusiva nas primeiras fases da vida da cartilagem, pois à medida que a matriz vai ganhando rigidez, a única forma de crescimento viável passa a ser por aposição.
De que forma ocorre o processo de reparo de uma cartilagem?
A cartilagem apresenta capacidade de tolerar níveis consideráveis e repetitivos de estresse. Porém, uma vez danificada, sua capacidade de cicatrizar é quase nula, mesmo em casos de lesões leves. Isso se deve a três motivos principais:

Ausência de capilares sanguíneos
Imobilidade
Baixa capacidade proliferativa de condrócitos
Algum nível de reparo até pode ocorrer, mas apenas quando o pericôndrio é o único atingido. Nesses casos, os agentes de reparo são as células progenitoras pluripotentes do pericôndrio, mas mesmo assim há baixa produção de células cartilaginosas. O reparo culmina principalmente na produção e deposição de tecido conjuntivo denso.
Tipos de cartilagem e formas de crescimento
A especialista Gabriela Cardoso Caldas faz uma revisão dos tipos de cartilagens e explica a osteoartrite, o tipo mais comum de doença articular.
Verificando o aprendizado
ATENÇÃO!
Para desbloquear o próximo módulo, é necessário que você responda corretamente a uma das seguintes questões:
O conteúdo ainda não acabou.
Clique aqui e retorne para saber como desbloquear.
MÓDULO 2
Descrever as características morfológicas do tecido ósseo, os componentes fundamentais, a estrutura geral e a formação dos diferentes tipos de ossos
Estrutura geral dos ossos e funções do tecido ósseo
O tecido ósseo é uma especialização do tecido conjuntivo que se caracteriza por possuir matriz extracelular mineralizada, constituída majoritariamente por fosfato de cálcio na forma de cristais de hidroxiapatita. A associação desses cristais a fibras colágenas é responsável pela rigidez e resistência característica do tecido ósseo, que proporciona suporte aos tecidos moles e proteção aos órgãos vitais, como aqueles contidos na caixa torácica.

Funções do tecido ósseo
São funções do tecido ósseo:
Armazenar cálcio, fosfato e outros íons
Por conta do seu conteúdo mineral, o osso também atua como um local de armazenamento para o cálcio, fosfato e outros íons, que podem ser liberados de maneira controlada, mantendo seus níveis adequados no organismo.
Abrigar e proteger órgãos vitais
O osso abriga e protege órgãos vitais ao organismo, como, por exemplo, a medula óssea, local de produção das células sanguíneas.
Promover a movimentação
O tecido ósseo é o principal componente do esqueleto. Fornece apoio aos músculos esqueléticos e forma um sistema de alavancas, que amplia as forças geradas na contração muscular.
Armazenar substâncias
Outra propriedade funcional dos ossos é a capacidade de absorver toxinas e metais pesados, minimizando as consequências maléficas em outros tecidos.
Em geral, um osso é constituído por outros tecidos conjuntivos além do tecido ósseo, como o tecido adiposo e tecido hematopoiético, além de vasos sanguíneos e nervos. A cartilagem hialina também é encontrada em situações nas quais o osso forma uma articulação livremente móvel com outro, a articulação sinovial.
Estrutura do osso
Agora que você já entendeu as muitas funções desempenhadas pelo tecido ósseo, é necessário compreender a estrutura do osso. Para isso, observe a figura a seguir.

Quando um osso é cortado, é possível identificar dois arranjos estruturais distintos:
- O primeiro é visualizado na parte externa como uma camada densa e compacta, que é denominada osso compacto.
- O segundo, na parte mais interna, consiste em uma rede semelhante a uma esponja, formada por trabéculas, denominada osso esponjoso.
Os espaços que você vê dentro da rede são contínuos e é onde encontramos a medula óssea (no canal medular) e os vasos sanguíneos. A localização do osso esponjoso e do osso compacto varia de acordo com o formato do osso. Dessa forma, podemos dividir os ossos em quatro grupos:

Ossos longos
Apresentam um corpo longo, denominado diáfise, e duas extremidades expandidas, as epífises. Nas epífises, podemos encontrar cartilagem hialina recobrindo a superfície articular. A porção alargada observada entre diáfise e epífise é chamada de metáfise e se estende da diáfise até a linha epifisária. Observando a porção interna, podemos perceber uma grande cavidade, que é ocupada pela medula óssea. Apenas uma pequena quantidade de osso esponjoso está voltada para a cavidade medular, já o osso compacto forma quase toda a espessura da diáfise. Já nas epífises, observa-se o inverso: a região de osso esponjoso é extensa enquanto o osso compacto consiste apenas em uma casca externa fina.
A tíbia e os ossos metacarpais (são os ossos que formam a primeira parte dos dedos das mãos) são exemplos de ossos longos.


Ossos curtos
Possuem comprimento e diâmetro quase iguais, geralmente formam articulações móveis com ossos vizinhos. Os ossos curtos, como os ossos carpais, apresentam uma camada exterior de osso compacto e osso esponjoso com o espaço medular em seu interior. A superfície externa do osso é recoberta por uma cápsula de tecido conjuntivo fibroso, o periósteo, que trataremos com mais detalhes adiante. Assim como os ossos longos, suas superfícies articulares são cobertas por cartilagem hialina.

Ossos planos ou chatos
Apresentam duas camadas espessas de osso compacto, com uma camada intermediária de osso esponjoso (díploe). Os ossos do crânio, como o frontal e occipital, e o esterno são exemplos de ossos planos: finos e semelhantes a placas.

Ossos irregulares
Possuem formato que não se encaixa em nenhum dos três grupos descritos acima, como as vértebras, que possuem morfologia complexa, e o etmoide, que contém espaços aéreos.
Etmoide
Localizado no centro do crânio, o etmoide contribui para os limites da órbita, da cavidade nasal, do septo nasal e do assoalho da fossa craniana anterior.
Vimos que a superfície externa dos ossos é recoberta por uma camada de tecido conjuntivo fibroso, o periósteo. As exceções são as áreas que se articulam com outro osso, que são cobertas por cartilagem. Na figura a seguir, vemos um corte transversal do osso, onde podemos observar áreas recobertas com periósteo e uma região com a cartilagem articular.

O periósteo consiste em uma camada fibrosa externa, que possui fibroblastos e fibras colágenas que se dispõem paralelamente à superfície do osso, formando uma cápsula e uma camada interna, que contém algumas poucas células osteoprogenitoras. Com estímulo apropriado, essas células são capazes de sofrer divisão e originar os osteoblastos.
Osteoblastos
Células secretoras de matriz óssea.
Você sabia
Chamamos de fibras de Sharpey os feixes de fibras colágenas do periósteo que penetram o tecido ósseo e prendem firmemente o periósteo ao osso.
A superfície interna do osso compacto, próxima à cavidade medular, e as trabéculas do osso esponjoso, como mostrado na figura a seguir, são revestidas pelo endósteo, que é composto frequentemente por apenas uma camada celular, com células osteoprogenitoras e células de revestimento ósseo. Essas células são similares, apresentando morfologia achatada e núcleo alongado. Por se localizarem no interior das cavidades ósseas, são também chamadas de células endosteais.

Como comentamos anteriormente, as trabéculas contínuas dos ossos esponjosos são ocupadas por vasos sanguíneos e pela medula óssea. A medula óssea vermelha consiste em células sanguíneas em diferentes estágios e uma rede estrutural de células articulares e fibras. Curiosamente, durante o crescimento do indivíduo, a quantidade de medula óssea não aumenta proporcionalmente ao crescimento do osso. Ela é geralmente limitada a espaços de osso esponjoso específicos, como o esterno e a crista ilíaca.
Em adultos, a velocidade de formação das células sanguíneas diminui e o tecido presente na cavidade medular consiste principalmente em adipócitos, sendo denominado medula amarela.
Medula amarela
Pode reverter-se em medula vermelha com estímulos apropriados, como uma extensa perda de sangue.
Características do tecido ósseo
Além das células osteoprogenitoras e das células de revestimento (periosteais e endosteais) que acabamos de estudar, outros tipos de célula também compõem o tecido ósseo. A seguir, abordaremos cada um deles.
Osteoblastos
Os osteoblastos são as células que apresentam grande capacidade de síntese e secretam os componentes orgânicos da matriz óssea, como colágeno tipo I, proteoglicanos e glicoproteínas. Sintetizam também a osteonectina, que facilita a deposição de cálcio na matriz, e a osteocalcina, que estimula a atividade dos osteoblastos. Além disso, participam da mineralização da matriz, pois são capazes de concentrar o fosfato de cálcio, como observado a seguir.
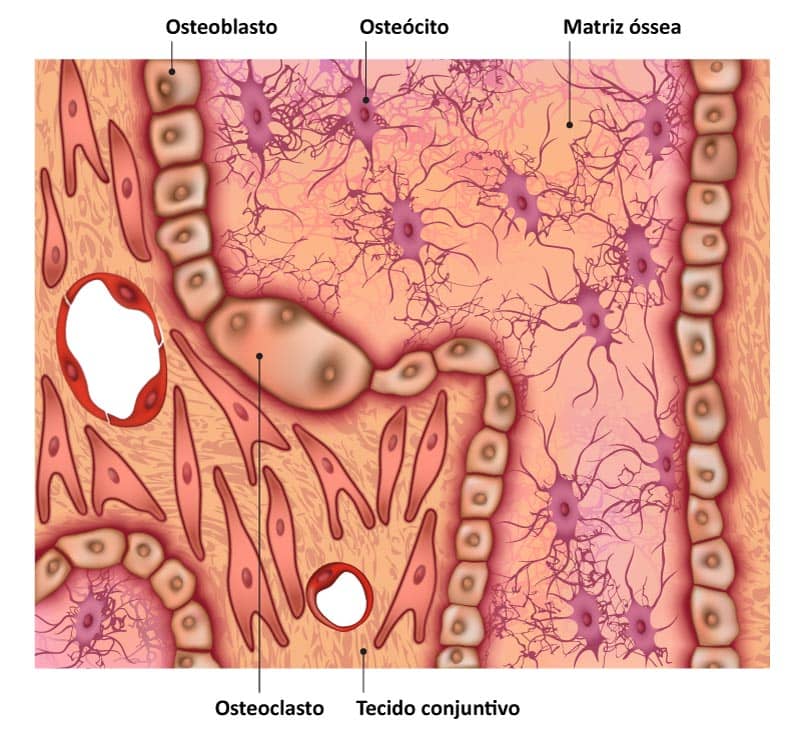
São encontrados nas superfícies ósseas, dispostos lado a lado, semelhante a um epitélio simples. Quando estão em atividade de síntese, apresentam morfologia cuboide e citoplasma basofílico. Porém, tornam-se achatados e reduzem a basofilia em estado pouco ativo. Durante a síntese e deposição da matriz ao redor da célula e de seus prolongamentos, ocorre a formação de lacunas e canalículos. Uma vez aprisionados por essa matriz óssea recém-sintetizada, os osteoblastos passam a ser chamados de osteócitos.
Osteócitos
Como vimos, os osteócitos são as células que ocupam as lacunas da matriz óssea, das quais partem os canalículos que possibilitam as trocas de nutrientes e substâncias entre essas células e os capilares sanguíneos. Cada lacuna é ocupada por apenas um osteócito, que é capaz de trocar pequenas moléculas e íons com outro osteócito por meio da existência de junções comunicantes em seus prolongamentos.
Apresentam morfologia achatada, retículo endoplasmático rugoso pouco abundante, cromatina nuclear condensada e complexo de Golgi pouco desenvolvido. Apesar de essas características indicarem baixa atividade de síntese, os osteócitos são células fundamentais para a manutenção da matriz óssea e sua morte é seguida pela reabsorção da matriz. Note os prolongamentos celulares (seta vermelha) dos osteócitos na figura a seguir.

Osteoclastos
Os osteoclastos são células de reabsorção óssea encontradas nas superfícies ósseas onde há remodelamento, remoção ou dano ósseo. São células gigantes e móveis, apresentando ramificações extensas e irregulares. Apresentam citoplasma granuloso, algumas vezes vacuolizado, basófilo nas células jovens e acidófilo nas maduras. A partir de mecanismos de regulação coordenados, os osteoclastos secretam colagenase, hidrolases e ácido (H+) nos ambientes de reabsorção óssea, digerindo localmente a matriz orgânica e dissolvendo o fosfato de cálcio. A origem dos osteoclastos se dá a partir de precursores mononucleados da medula óssea, que quando entram em contato com o tecido ósseo, unem-se e formam os osteoclastos, multinucleados, como mostrado na figura a seguir.
Colagenase
Enzimas que quebram as ligações peptídicas presentes na molécula de colágeno.
Hidrolases
Enzimas que promovem a quebra de substâncias orgânicas a partir da utilização de moléculas de água.

De fato, a parte inorgânica representa cerca de 50% do peso da matriz. A parte orgânica e a inorgânica apresentam algumas peculiaridades, vamos conhecê-las?
Parte inorgânica da matriz
O fosfato e o cálcio são os íons mais abundantes, mas também podemos encontrar bicarbonato, magnésio, potássio, sódio e citrato em pequenas quantidades.
Parte orgânica da matriz
Como já comentamos brevemente, é constituída majoritariamente (95%) por fibras colágenas do tipo I e, em menor quantidade, por glicoproteínas e proteoglicanos. Um fato curioso é que as glicoproteínas do osso podem participar em algum nível da mineralização da matriz, uma vez que outros tecidos, também ricos em colágeno do tipo I, mas que não possuem essas glicoproteínas, normalmente não se calcificam.
Na figura a seguir, vemos um resumo de todas as células associadas aos ossos. Note a origem dos osteoblastos a partir de células-troncos mesenquimais e osteoclastos a partir do progenitor de granulócitos/monócitos.

Você sabia
A matriz óssea recém-sintetizada, ainda não calcificada, próxima aos osteoblastos ativos recebe o nome de osteoide.
Tipos de tecido ósseo
Do ponto de vista histológico, existem dois tipos de tecido ósseo: o primário ou imaturo e o secundário, maduro ou lamelar. Ambos contêm os mesmos constituintes celulares e de matriz, mas apresentam diferenças no período de existência e organização das fibras colágenas.
Tecido ósseo primário ou imaturo
O tecido ósseo primário é o que aparece primeiro em cada osso, tanto no desenvolvimento embrionário quanto no reparo de fraturas, sendo gradualmente substituído pelo tecido ósseo secundário. É pouco frequente em indivíduos adultos, encontrado apenas próximo às suturas dos ossos do crânio, nos alvéolos dentários e em alguns pontos de inserção de tendões.
Atenção
É o osso imaturo nos alvéolos dentários que possibilita as tão difundidas correções ortodônticas. O tecido primário apresenta menor quantidade de minerais, maior proporção de osteócitos e fibras colágenas dispostas em várias direções, de maneira desorganizada.
Note, na figura a seguir, a quantidade de osteócitos (núcleos celulares são os pontos roxos) e fibras colágenas dispostas sem orientação definida.

Tecido ósseo secundário, maduro ou lamelar
A principal característica do tecido ósseo secundário é a existência de fibras colágenas organizadas em lamelas, que ficam paralelas umas às outras ou em camadas concêntricas em torno de canais com vasos sanguíneos, formando o que chamamos de ósteons ou sistemas de Havers. Entre os grupos de lamelas, observa-se frequentemente o acúmulo de uma matriz mineralizada pobre em colágeno. As lacunas que abrigam os osteócitos geralmente se situam entre as lamelas, mas algumas vezes estão dentro delas. Note, na figura a seguir, os ósteons representados na figura pelo símbolo “O” e as camadas de lamelas concêntricas.

Cada sistema de Havers é formado por 4 a 20 lamelas ósseas concêntricas, constituindo um cilindro longo, às vezes bifurcado, paralelo à diáfise. No centro desse cilindro existe o canal de Havers, revestido pelo endósteo, contendo nervos e vasos sanguíneos. A seguir conseguimos observar um esquema ilustrando a estrutura de um ósteon.

Se fizermos cortes transversais à diáfise, enxergaremos facilmente uma organização típica das lamelas ósseas constituindo os sistemas de Havers: circunferenciais interno, externo e os intermediários. Os sistemas de Havers são característicos da diáfise dos ossos longos, mas sistemas pequenos também podem ser encontrados no osso compacto de outros locais. A comunicação dos canais de Havers, entre si, com a cavidade medular e com a superfície externa se dá por canais oblíquos ou transversais que atravessam as lamelas ósseas: os canais de Volkmann.
Canais de Volkmann
Ao contrário dos canais de Havers, não apresentam lamelas ósseas concêntricas.
Todos os canais vasculares do tecido ósseo aparecem quando a matriz se forma ao redor dos vasos preexistentes.
Formação e mineralização óssea
O tecido ósseo pode ser formado por dois processos:
Ossificação endocondral
No qual um molde de cartilagem hialina serve de precursor do osso. São exemplos de ossos que se desenvolvem a partir desse processo os ossos dos membros inferiores e superiores, assim como as partes do esqueleto que sustentam peso (vértebras, por exemplo).
Ossificação intramembranosa
Processo mais simples que ocorre no interior de uma membrana conjuntiva. Desenvolvem-se por esse processo os ossos chatos do crânio, da face, a mandíbula e a clavícula.
Atenção
Em ambos os processos, forma-se primeiro o osso primário, que é gradualmente substituído pelo secundário. Apesar de existirem esses dois tipos de ossificação, isso não significa que determinado osso seja classificado como membranoso ou endocondral. Esses termos referem-se apenas ao processo inicial de formação óssea.
Por conta da remodelação que ocorre em seguida, o tecido ósseo inicialmente depositado por algum desses dois mecanismos é posteriormente substituído. O osso de reposição é formado sobre o osso que já existe por crescimento aposicional.
Portanto, durante o crescimento dos ossos, podemos encontrar áreas de tecido primário, áreas de absorção e áreas de tecido secundário. A combinação de formação e remodelamento ósseo persiste durante o crescimento do osso, mesmo que em um ritmo bem mais lento em adultos.
Agora vamos entender melhor esses dois processos.
Ossificação intramembranosa
Como você já sabe, a ossificação intramembranosa ocorre no interior de membranas de tecido conjuntivo, local chamado de centro de ossificação primária. Inicialmente, há a diferenciação das células mesenquimais em osteoblastos, que sintetizam a matriz ainda não mineralizada. Logo em seguida, há a mineralização da matriz, que engloba os osteoblastos e esses, por sua vez, se transformam em osteócitos.
Durante esse processo, há a confluência de várias traves de matriz, dando ao osso um aspecto esponjoso. As cavidades formadas são penetradas por vasos sanguíneos e pelas células mesenquimatosas que darão origem à medula óssea. A seguir, apresentamos as etapas da ossificação intramembranosa:
Primeira etapa
Surge um centro de ossificação no tecido conjuntivo mesenquimatoso. Ocorre diferenciação das células osteoprogenitoras derivadas do mesênquima em osteoblastos. Os osteoblastos começam a secretar a matriz não mineralizada, o osteoide.


Segunda etapa
Os osteoblastos acumulam-se na periferia do centro de ossificação e continuam secretando osteoide em direção ao centro de ossificação. À medida que o processo continua, a matriz se mineraliza e engloba os osteoblastos, que se diferenciam em osteócitos.
Terceira etapa
O tecido recém-formado tem uma estrutura microscópica de osso imaturo com trabéculas espessas revestidas por osteoblastos e células endosteais.


Quarta etapa
O crescimento adicional e a remodelação do osso resultam na substituição do osso imaturo por um osso maduro, compacto e lamelar. Entre as lamelas de osso compacto há trabéculas de osso esponjoso. Os espaços entre as trabéculas são ocupados por células da medula óssea, conduzidas por vasos sanguíneos.
Os vários centros de ossificação crescem radialmente e, então, substituem a membrana conjuntiva preexistente. O crescimento adicional por aposição e a remodelação resultam em substituição do osso primário por osso compacto na periferia da peça óssea e por osso esponjoso no centro do osso recém-formado. Os ossos frontal e parietal, assim como partes dos ossos occipital, temporal e dos maxilares, são inicialmente formados por ossificação intramembranosa, que também contribui para o crescimento dos ossos curtos e aumento da espessura dos ossos longos. Nos ossos chatos do crânio, por exemplo, a parte da membrana conjuntiva que não sofre calcificação constitui o endósteo e periósteo.
Você sabia
Durante a palpação do crânio de recém-nascidos, podemos sentir as fontanelas, áreas moles onde as membranas conjuntivas ainda não foram substituídas por tecido ósseo.
Ossificação endocondral
A ossificação endocondral, como já comentamos, usa um molde de cartilagem hialina como precursor para o desenvolvimento ósseo. Primeiramente, ocorrem modificações na cartilagem que envolvem hipertrofia de condrócitos, redução da matriz cartilaginosa a finos tabiques, mineralização e apoptose dos condrócitos.
Posteriormente, as cavidades que eram ocupadas pelos condrócitos são invadidas por capilares sanguíneos e células osteogênicas do tecido conjuntivo vizinho, que se diferenciam em osteoblastos e começam a secretar matriz sobre os tabiques de cartilagem calcificada. Como você pode perceber, não há transformação do tecido cartilaginoso em tecido ósseo, mas o aparecimento do tecido ósseo onde antes havia cartilagem. Os tabiques da matriz cartilaginosa servem como pontos de suporte para que a ossificação ocorra.
A ossificação endocondral é a principal responsável pela formação dos ossos curtos e longos. A formação dos ossos longos é um processo complexo, que estudaremos com mais detalhes a seguir.
O molde cartilaginoso dos ossos longos apresenta uma parte média estreita e extremidades dilatadas, que correspondem à diáfise a às epífises futuras. O primeiro tecido ósseo que aparece nos ossos longos é formado por ossificação intramembranosa do pericôndrio, que recobre parte da diáfise e forma uma espécie de cilindro chamado de colar ósseo.
Durante o desenvolvimento do colar ósseo, as células cartilaginosas envolvidas por ele sofrem hipertrofia, morrendo por apoptose, e a matriz da cartilagem se mineraliza. A partir daí, vasos sanguíneos vindos do periósteo atravessam o colar ósseo e penetram na cartilagem recém-calcificada, levando células osteoprogenitoras. Com a proliferação dessas células e diferenciação em osteoblastos, há a formação de camadas celulares na superfície dos tabiques cartilaginosos e secreção da matriz óssea que rapidamente se mineraliza. Observe os detalhes dessa formação a seguir.

Saiba mais
Na análise histológica, a cartilagem calcificada é basófila, diferentemente do tecido ósseo, que é acidófilo.
O centro de ossificação é chamado de centro primário, que cresce rapidamente e ocupa toda a diáfise. Esse alastramento é acompanhado pelo cilindro ósseo, que cresce também em direção às epífises. Desde o início da formação do centro primário, há o aparecimento de osteoclastos e absorção do tecido ósseo, formando o canal medular.
À medida que o canal medular se desenvolve e também cresce longitudinalmente, as células-tronco hematopoiéticas trazidas pelo sangue dão origem à medula óssea, que será responsável pela produção de todas as células sanguíneas, tanto na vida intrauterina como após o nascimento do indivíduo.
E as epífises?
Um pouco mais tarde, formam-se os centros secundários de ossificação, um em cada epífise. Eles não aparecem simultaneamente e são semelhantes aos centros primários, mas possuem crescimento radial. Também encontramos medula óssea na porção central das epífises.
Por fim, o tecido cartilaginoso fica reduzido a dois locais:
Cartilagem articular
Persistirá por toda a vida e não contribui para a formação de tecido ósseo.
Disco epifisário ou cartilagem de conjugação
Encontrada entre as epífises e a diáfise e que, a partir desse momento, torna-se responsável pelo crescimento longitudinal do osso até os 20 anos de idade do indivíduo, quando desaparece por ossificação.

Sobre o mecanismo de calcificação da matriz, ainda não existe uma hipótese que seja universalmente aceita. O que sabemos é que a calcificação começa com a deposição de sais de cálcio sobre as fibras de colágeno, processo que parece ser induzido pelos proteoglicanos e pelas proteínas de matriz. Além disso, a deposição sofre influência da enzima fosfatase alcalina, sintetizada pelos osteoblastos, e da concentração de sais de cálcio em vesículas dessas células, que são expelidas para a matriz.
Articulações
Você já parou para pensar como os ossos se ligam uns aos outros para constituir o nosso esqueleto?
Quem promove essa união são estruturas formadas por tecidos conjuntivos, as articulações. Elas podem ser classificadas em: sinartroses e diartroses.
As sinartroses não possibilitam movimentos, ou quando ocorrem, são muito limitados. Podem ser divididas em três tipos, dependendo do tecido que une os ossos:
Os ossos são unidos por tecido ósseo, e as articulações são totalmente ausentes de movimentos. São encontradas unindo os ossos chatos do crânio em idosos. Em adultos jovens e crianças, a união é dada por tecido conjuntivo denso.
Os ossos são unidos por cartilagem hialina e as articulações apresentam movimentos limitados. São encontradas na articulação da primeira costela com o esterno.
Os ossos são unidos por tecido conjuntivo denso e, assim como as sincondroses, são dotadas de algum movimento. A sínfise pubiana e a articulação tibiofibular inferior são exemplos de sindesmoses.
Já as diartroses são articulações que possuem grande mobilidade e geralmente unem os ossos longos. Observando a figura a seguir, podemos notar uma cápsula que liga as extremidades ósseas e delimita uma cavidade fechada, a cavidade articular. A cápsula é formada por duas partes: a camada fibrosa externa e a camada sinovial (membrana sinovial) que reveste a cavidade articular, exceto as áreas de cartilagem.

A cavidade articular, como mostrada na figura ao lado, contém um líquido viscoso e transparente, o líquido sinovial. Esse líquido é um dialisado do plasma sanguíneo com alto teor de ácido hialurônico, produzido pelas células da camada sinovial.
Você deve lembrar que as superfícies articulares que são revestidas por cartilagem hialina não apresentam pericôndrio e, portanto, o seu deslizamento é facilitado pelo efeito lubrificante do ácido hialurônico. Além disso, o líquido sinovial funciona como uma via de transporte de substâncias entre a cartilagem articular, que é avascular, e os capilares sanguíneos da membrana sinovial. Enquanto nutrientes e oxigênio passam do sangue para a cartilagem, o CO2 faz o caminho inverso.

As diartroses possuem cápsulas com estruturas diferentes, conforme a articulação. De maneira geral, as cápsulas são constituídas por uma camada externa, a camada fibrosa, e uma camada interna, a membrana ou camada sinovial.
Revestindo a camada sinovial, encontramos dois tipos celulares: um semelhante ao fibroblasto e o outro com morfologia e atividade funcional semelhantes ao macrófago. A camada fibrosa da cápsula articular é formada por tecido conjuntivo denso.
Osteoporose e fatores nutricionais na formação óssea
A especialista Gabriela Cardoso Caldas explica sobre o impacto da nutrição na formação óssea e sobre a osteoporose, a principal doença óssea.
Verificando o aprendizado
ATENÇÃO!
Para desbloquear o próximo módulo, é necessário que você responda corretamente a uma das seguintes questões:
O conteúdo ainda não acabou.
Clique aqui e retorne para saber como desbloquear.
MÓDULO 3
Reconhecer os três tipos de tecido muscular, as suas características morfológicas e as diferentes formas de contração muscular
Visão geral, classificação e função dos músculos
O tecido muscular é um dos quatro tipos básicos de tecido do organismo. Ele é responsável pelo movimento dos membros, do corpo como um todo e pelas mudanças no tamanho e formato dos órgãos internos. Caracteriza-se pela presença de agregados de células alongadas, especializadas e dispostas em arranjos paralelos. Essas células possuem grande quantidade de filamentos citoplasmáticos de proteínas contráteis, que geram forças necessárias para a contração desse tecido a partir da hidrólise de moléculas de adenosina trifosfato (ATP).
Saiba mais
A adenosina trifosfato (ATP) é um nucleotídeo formado por uma ribose (açúcar) ligada à adenina (base nitrogenada) e três grupos fosfato em série. É uma molécula que armazena energia, liberando-a, quando necessário, por uma reação de hidrólise (em que ocorre a quebra de uma molécula em moléculas menores com a participação da água). Nessa reação, são formados adenosina difosfato (ADP), fosfato inorgânico (Pi) e energia.
As células musculares se originam no mesoderma, e sua diferenciação ocorre pela síntese das proteínas filamentosas, concomitante ao alongamento celular. Elas são células alongadas e, por isso, são também chamadas fibras musculares. Sua matriz extracelular consiste na lâmina basal e nas fibras reticulares.
De acordo com as características morfológicas e funcionais, o tecido muscular pode ser dividido em três tipos:

Músculo estriado esquelético
Está inserido nos ossos e é responsável pelo movimento do esqueleto, bem como pela posição e postura corporal. Nos olhos, esse tipo de músculo possibilita o preciso movimento dos olhos. É constituído por feixes de células cilíndricas muito longas e multinucleadas, que apresentam estrias transversais. Essas células, também chamadas de fibras, apresentam contração rápida e vigorosa, com controle voluntário.

Músculo estriado cardíaco
É encontrado na parede do coração e em pequenos trechos das grandes veias pulmonares que desembocam no coração. Suas células, alongadas e ramificadas, também apresentam estriações transversais e se unem por meio dos discos intercalares, estruturas exclusivas desse tipo de músculo. As células cardíacas, ou cardiomiócitos, apresentam contração involuntária, rítmica e vigorosa.

Músculo liso
É encontrado nas vísceras, no sistema circulatório, nos músculos intrínsecos do olho e nos músculos que fazem os pelos da pele se levantarem. É formado por aglomerados de células fusiformes, que não apresentam estrias transversais. Sua contração é lenta e está sob controle involuntário.
Você sabia
Nas células musculares, a membrana celular é chamada de sarcolema, o citosol, de sarcoplasma, e o retículo endoplasmático liso, de retículo sarcoplasmático.
Veja a seguir um esquema simplificado da fibra muscular.

Agora que você foi apresentado às características gerais do tecido muscular, conheceremos com mais detalhes cada um dos tipos de tecido citados.
Músculo estriado – esquelético
Como já comentamos, o músculo esquelético é formado por feixes de células alongadas (que podem chegar a até 30cm e diâmetro de 10 a 100µm), cilíndricas, multinucleadas e com muitos filamentos, as miofibrilas. As fibras esqueléticas se originam na fase embrionária pela fusão dos mioblastos, que também são células alongadas. Os numerosos núcleos possuem localização periférica, próximos ao sarcolema. A localização do núcleo ajuda na distinção para as fibras cardíacas, onde os núcleos são centrais. Note na figura a seguir as estrias transversais.

Saiba mais
As células musculares esqueléticas do adulto não sofrem mitoses, mas é possível a formação de novas células no processo de reparo após lesão ou de hipertrofia decorrente do exercício intenso, pela divisão e fusão de mioblastos quiescentes, as células satélites.
Organização muscular
Em um músculo, as fibras se organizam em grupos de feixes, sendo o conjunto envolvido por uma camada de tecido conjuntivo, o epimísio, que recobre o músculo inteiro. Do epimísio partem finos septos do conjuntivo, o perimísio, que entram no músculo e separam os feixes. Dessa forma, o perimísio envolve os feixes de fibras (fascículo muscular). Por fim, cada fibra muscular é envolvida individualmente pelo endomísio, formado pela lâmina basal da fibra muscular associada a fibras reticulares.
Observe, na figura a seguir, os detalhes dessa organização muscular.

As fibras musculares são mantidas unidas pelo tecido conjuntivo, possibilitando que a força de contração gerada individualmente por cada fibra atue no músculo inteiro. Esse papel é de extrema importância funcional, uma vez que a maioria das fibras não alcança as duas extremidades musculares. Além disso, por meio do tecido conjuntivo, a força da contração do músculo é transmitida para outras estruturas, como os tendões e ossos. A força da contração pode ser regulada a partir do número de fibras musculares estimuladas pelos nervos.
Dos septos do conjuntivo partem os vasos sanguíneos que penetram os músculos e formam uma extensa rede de capilares entre as fibras, além de vasos linfáticos e nervos.
Organização das fibras
As fibras musculares esqueléticas apresentam estrias transversais, pela alternância de faixas claras e escuras. Por conta das características que apresentam ao microscópio de polarização, as faixas escuras são denominadas de banda A, e as faixas claras, de banda I. No centro das bandas I existe uma linha transversal escura, a linha Z, e no centro das bandas A existe uma zona mais clara, a banda H. Essas bandas formam o sistema de estriações transversais, paralelas, característico das fibras esqueléticas. Para entender melhor esses termos, observe a figura a seguir.

Cada fibra muscular contém diversos feixes cilíndricos de filamentos, as miofibrilas. A estriação da miofibrila deve-se à repetição dos sarcômeros, que são unidades iguais formadas pela parte da miofibrila que fica entre duas linhas Z sucessivas e contém duas semibandas I separadas por uma banda A, conforme observamos na figura a seguir.

Dispostos longitudinalmente nas miofibrilas, encontramos filamentos finos, de actina e outras proteínas, e grossos, de miosina. Esses filamentos miofibrilares, observados ao microscópio eletrônico de transmissão, distribuem-se de maneira simétrica e paralela, disposição que é mantida por várias proteínas. Uma dessas proteínas é a desmina, cujos filamentos ligam as miofibrilas umas às outras. O conjunto de miofibrilas é preso à membrana plasmática da célula muscular por meio de outras várias proteínas, como a distrofina.
Os filamentos finos partem da linha Z e vão até a borda externa da banda H. Já os filamentos de miosina ocupam a região central do sarcômero. Dessa forma:
| Quadro Resumo | |
|---|---|
| Banda I | Formada somente por filamentos finos. |
| Banda A | Formada pelos dois tipos de filamentos. |
| Banda H | Formada somente por filamentos de miosina. |
Saiba mais
Na banda A, os filamentos de miosina são rodeados por seis filamentos finos, formando um hexágono.
Veja, na figura a seguir, como é esse aspecto do sarcômero.

A miosina e a actina representam mais da metade das proteínas do músculo estriado. No entanto, além da actina e da miosina, as miofibrilas do músculo estriado contêm outras duas proteínas principais, a tropomiosina e a troponina.
Observe, na figura a seguir, que cada molécula de tropomiosina (feixe em amarelo) ocupa o sulco que corresponde a sete moléculas de actina (representada pelas bolas rosas) e, para cada molécula de tropomiosina, existe um complexo de troponina. Quando há o estímulo muscular, a molécula de troponina altera seu formato, afunda a tropomiosina no sulco e libera os sítios da actina de interação com a miosina.

Agora que você está familiarizado com a organização e composição das fibras musculares, chegou o momento de entender como ocorre o mecanismo de contração muscular.
Mecanismos de contração muscular
No sarcômero em repouso, os filamentos finos e grossos se sobrepõem parcialmente e, durante a contração, mantêm seus comprimentos originais. O que ocorre durante a contração é o deslizamento dos filamentos uns sobre os outros, aumentando o tamanho da região de sobreposição e diminuindo o tamanho do sarcômero.
Por quê?
Uma vez que os filamentos de actina penetram na banda A, a banda I diminui de tamanho. Ao mesmo tempo, a banda H também se reduz, à medida que os filamentos finos se sobrepõem completamente aos grossos. Como resultado, cada sarcômero e, em consequência, a fibra muscular inteira sofrem encurtamento.
Achou confuso?
A figura a seguir apresenta como isso acontece.

A contração muscular é um evento que depende da disponibilidade de íons cálcio (Ca2+), armazenados no retículo sarcoplasmático, e é responsável pelo controle de nervos motores, que se ramificam no tecido conjuntivo do perimísio. O local de contato com a fibra muscular é denominado de placa motora ou junção mioneural. Quando a fibra do nervo motor recebe um impulso nervoso (potencial de ação), seu terminal axônico libera acetilcolina, um neurotransmissor que se difunde pela fenda sináptica e se associa a receptores do sarcolema. A ligação da acetilcolina resulta em uma maior permeabilidade da membrana ao sódio, despolarizando o sarcolema e, em seguida, o retículo sarcoplasmático.
Impulso nervoso
É uma rápida alteração de potencial elétrico das membranas, que possibilita a condução de sinais elétricos. As células que geram e conduzem os potenciais de ação possuem uma membrana excitável. Quando em repouso, o meio intracelular apresenta carga global negativa e o extracelular, positiva. No início do impulso nervoso, há uma despolarização da membrana, ou seja, uma inversão das cargas que permite a transmissão do sinal.
Quando a membrana do retículo é despolarizada por estímulos nervosos, os canais de cálcio se abrem, e esses íons, que antes estavam armazenados em cisternas, difundem-se passivamente (sem gasto de energia) e atuam sobre a troponina. Uma vez cessada a despolarização, a membrana do retículo transfere os íons de cálcio para o interior das suas cisternas, em um processo ativo (com consumo de energia) que interrompe a contração.
Veja a seguir o estímulo nervoso na fibra muscular esquelética.

O processo de contração no músculo estriado envolve uma série de eventos bioquímicos e mecânicos. A miosina, cuja cabeça tem atividade ATPase, converte a energia química em força mecânica a partir dos estados de associação e dissociação com a actina.
Durante o repouso, uma molécula de ATP está ligada à cabeça da miosina. Para que ocorra a hidrólise do ATP e a liberação de energia, a miosina precisa da actina como cofator. Quando o músculo está em repouso, a interação miosina-actina não ocorre devido à presença do complexo troponina-tropomiosina sobre os filamentos de actina, como observamos na figura a seguir.

Porém, quando há disponibilidade de íons de cálcio, estes se ligam a uma das três subunidades da troponina, mudando a configuração do complexo das três subunidades. Com a mudança estrutural da troponina, a molécula de tropomiosina é empurrada mais para dentro do sulco de actina. O resultado é que os sítios de ligação da actina com a miosina são expostos, propiciando a interação entre as cabeças de miosina com a actina. Dessa forma, o ATP libera adenosina difosfato (ADP), fosfato inorgânico e energia.
Com a hidrólise do ATP, ocorre uma ligeira deformação na miosina, aumentando a curvatura de sua cabeça. Como a actina está interagindo com a miosina, o movimento da cabeça da miosina empurra o filamento de actina, promovendo seu deslizamento sobre o filamento de miosina. Observe essa interação na figura a seguir.

Embora o filamento grosso tenha várias cabeças de miosina, em cada momento da contração apenas um pequeno número alinha-se com os sítios de actina.
Veja na figura a seguir essa interação.

Conforme as cabeças de miosina movimentam a actina, surgem novos locais para formação de interações actina-miosina. As pontes antigas de actina-miosina somente se desfazem depois que a miosina se une a uma nova molécula de ATP, o que determina também o retorno da cabeça de miosina para sua posição inicial, pronta para um novo ciclo.
Saiba mais
Sem ATP, o complexo actina-miosina fica estável, o que explica a rigidez muscular que ocorre logo após a morte, o rigor mortis.
Como você deve ter notado, uma única contração muscular é resultado de milhares de ciclos de formação e destruição de interações actina-miosina. A atividade contrátil continua acontecendo até que os íons de cálcio sejam removidos e o complexo troponina-tropomiosina cubra novamente o sítio de interação da actina com a miosina.
Iniciada no sarcolema, a despolarização teria que se difundir por toda a espessura da fibra para alcançar as cisternas de cálcio do retículo mais profundas. Sabendo disso, você deve estar imaginando que, em fibras musculares de maior calibre, as miofibrilas periféricas se contrairiam antes das mais profundas, certo?
A garantia de que a contração de cada fibra muscular esquelética se dê de maneira uniforme é dada pelo sistema de túbulos transversais, ou sistema T. Esse sistema é constituído por uma rede de invaginações do sarcolema, cujos ramos envolvem as junções das bandas A e I de cada sarcômero. O sinal despolarizador passa para o retículo sarcoplasmático e resulta na liberação de cálcio, que inicia o ciclo de contração. Veja, na figura a seguir, um esquema desse processo.

Atenção
A destruição da acetilcolina é necessária para evitar o contato prolongado com seu receptor. Uma vez terminada a despolarização, o cálcio é transportado de volta para as cisternas do retículo, a contração cessa, e a fibra muscular relaxa.
Músculo estriado – cardíaco
O tecido muscular cardíaco é composto por células alongadas e ramificadas (10 a 20μm de diâmetro e 80 a 100μm de comprimento), aderidas umas às outras por meio de junções intercelulares complexas. Essas células também apresentam estrias transversais, mas ao contrário das células esqueléticas, apresentam somente um ou dois núcleos, dispostos de maneira central, como mostrado na figura ao lado. As fibras cardíacas são envoltas por uma delicada bainha de tecido conjuntivo, equivalente ao endomísio que comentamos anteriormente, com uma abundante rede de capilares sanguíneos.

Sabemos que uma característica exclusiva do músculo cardíaco é a presença de linhas transversais fortemente coráveis, denominadas discos intercalares, que cruzam as fibras musculares de modo linear ou em forma de escada. Observando os discos intercalares em forma de escada, distinguem-se duas regiões:
Parte transversal
Cruza a fibra em ângulo reto.
Parte lateral
Paralela aos miofilamentos.
Os discos intercalares dispõem-se de maneira irregular ao longo das fibras e representam regiões de fixação altamente especializadas entre células vizinhas, conforme observamos na figura a seguir:

Encontramos três especializações juncionais principais nos discos intercalares:
Representam a principal especialização de membrana celular da parte transversal do disco, são encontradas também nas partes laterais e servem para ancorar os filamentos de actina dos sarcômeros terminais.
Unem as células cardíacas, impossibilitando que elas se separem durante a atividade contrátil.
Encontradas nas partes laterais dos discos, são responsáveis pela comunicação iônica entre as células vizinhas. Com isso, o sinal iônico passa como uma onda de uma célula para outra.
Na imagem a seguir, conseguimos observar a disposição do disco intercalar e suas especializações juncionais.

As proteínas contráteis das células musculares cardíacas apresentam estrutura e função praticamente iguais às das células esqueléticas. Porém, no músculo cardíaco, o sistema T e o retículo sarcoplasmático não são tão bem organizados como no músculo esquelético. Os ventrículos apresentam sistemas T maiores que os dos músculos esqueléticos e que se localizam na altura da banda Z. Já o retículo sarcoplasmático apresenta distribuição irregular entre os miofilamentos e não é tão desenvolvido.
Você sabia
Enquanto nas fibras musculares esqueléticas o túbulo T apresenta conexão com duas cisternas do retículo sarcoplasmático (estrutura conhecida como tríade), nas fibras cardíacas, os túbulos T geralmente se associam a apenas uma cisterna, estrutura chamada de díade.
Outra característica das células musculares cardíacas é a presença de numerosas mitocôndrias. Elas ocupam aproximadamente 40% do volume celular, contra apenas 2% ocupado pelas mitocôndrias das células esqueléticas.
Essa quantidade abundante reflete o intenso metabolismo aeróbio realizado pelo tecido. As células musculares cardíacas armazenam ácidos graxos na forma de triglicerídeos, encontrados em gotículas lipídicas citoplasmáticas. Também há glicogênio, mas em pequena quantidade, fornecendo glicose quando há necessidade. Por fim, também encontramos no músculo cardíaco uma rede de células musculares modificadas, acopladas a outras células musculares, que apresentam função importante na geração e condução do estímulo cardíaco, de tal modo a cadenciar as contrações atriais e ventriculares.
Músculo liso
O músculo liso é composto por células longas, espessas no centro e mais delgadas nas extremidades. As células musculares lisas, também denominadas fibras, não apresentam o padrão estriado encontrado no músculo esquelético e no músculo cardíaco. O tamanho celular pode variar, desde 20μm na parede dos pequenos vasos sanguíneos até 500μm no útero de uma mulher grávida. Inclusive, durante a gravidez, as fibras musculares do útero aumentam de número (hiperplasia) e de tamanho (hipertrofia). Essas células são revestidas por lâmina basal e se mantêm unidas por uma rede de fibras reticulares, de modo que a contração simultânea de apenas algumas células reverbere pelo músculo inteiro. Observe, na figura a seguir, a ausência das estrias transversais no músculo liso.

Um aspecto particular das células musculares lisas é a existência de abundantes invaginações da membrana celular, semelhantes a cavéolas. Acredita-se que elas funcionem de forma análoga ao sistema de túbulos T que comentamos anteriormente, para liberação de Ca2+ no citoplasma. Os níveis intracelulares de Ca2+ também são muito importantes na regulação da contração do músculo liso.
Cavéolas
A membrana celular forma pequenas invaginações em forma de bolsa para o interior do citoplasma.
Além de algumas mitocôndrias, cisternas do retículo endoplasmático rugoso, grânulos de glicogênio e complexo de Golgi pouco desenvolvido, as células musculares lisas também apresentam corpos densos. Essas estruturas se localizam principalmente na membrana celular, podendo existir também no citoplasma, e possuem importante papel na contração das células musculares lisas (podemos compará-los às linhas Z dos músculos estriados). O mecanismo envolvido será abordado a seguir. A célula muscular lisa, além da sua capacidade contrátil, também pode sintetizar colágeno do tipo III (que forma as fibras reticulares), fibras elásticas e proteoglicanos.
Veja, na figura a seguir, o aspecto de uma célula muscular lisa relaxada e contraída.

No músculo liso, a contração também depende do deslizamento dos filamentos de actina e miosina, mas o mecanismo molecular envolvido é diferente do que aprendemos para os músculos estriados.
Encontramos no sarcoplasma das células musculares lisas filamentos de actina estabilizados pela combinação com a tropomiosina. Porém, aqui não existem sarcômeros nem troponina, e os filamentos de miosina só se formam no momento da contração.
Vimos que, no tecido muscular estriado, a miosina (que é do tipo I) existe permanentemente estirada, constituindo os filamentos grossos. Já nas células musculares lisas, encontramos a miosina II, cujas moléculas ficam enroladas. Uma vez combinadas com um radical fosfato, elas se distendem e formam um filamento.
A contração nas células musculares lisas ocorre da seguinte maneira:
- A partir do estímulo do sistema nervoso autônomo, os íons de cálcio migram do meio extracelular para o sarcoplasma pelos canais de transporte especializados localizados na membrana plasmática.
- Os íons de cálcio se combinam com uma proteína, a calmodulina, formando um complexo que ativa a enzima quinase da cadeia leve da miosina II.
- A enzima ativa fosforila as moléculas de miosina II, que se distendem e assumem a forma filamentosa. Com essa mudança conformacional, os sítios que possuem atividade ATPase e que se combinarão com a actina ficam descobertos.
- A combinação da miosina com a actina libera energia do ATP, que promove a deformação da cabeça da miosina II e, com isso, ocorre o deslizamento dos filamentos de actina e de miosina uns sobre os outros.
Sistema nervoso autônomo
É a porção do sistema nervoso central (SNC) que controla a maioria das funções viscerais do organismo.
A actina e miosina ligam-se a filamentos intermediários de desmina e de vimentina, que por sua vez prendem-se aos corpos densos da membrana celular. Essa configuração provoca a contração da célula como um todo.
Veja a seguir um esquema da contração da célula muscular lisa.

Além do cálcio, outros fatores ativam a miosina II e estimulam a contração celular. Um exemplo é o aumento nos níveis citosólicos de AMP-cíclico, que pode ser induzido por hormônios sexuais, como os estrogênios.
AMP-cíclico
É formada a partir do ATP, uma importante molécula na transdução de sinal, sendo responsável pela modulação de processos fisiológicos.
A inervação do músculo liso é realizada por fibras do sistema nervoso simpático e parassimpático, porém não há placas motoras como no músculo esquelético. Frequentemente, os axônios formam dilatações entre as células musculares lisas, que contêm vesículas sinápticas com os neurotransmissores acetilcolina ou norepinefrina.
Acetilcolina
Conhecidas como terminações colinérgicas.
Norepinefrina
Conhecidas como terminações adrenérgicas.
Algumas dessas dilatações axônicas estão bem próximas da célula muscular, porém há outras mais distantes. As terminações nervosas adrenérgicas e colinérgicas atuam de modo antagônico, estimulando ou inibindo a contração do músculo.
Músculo estriado esquelético: tipos de fibras
A especialista Gabriela Cardoso Caldas explica algumas características do músculo estriado esquelético.
Verificando o aprendizado
ATENÇÃO!
Para desbloquear o próximo módulo, é necessário que você responda corretamente a uma das seguintes questões:
O conteúdo ainda não acabou.
Clique aqui e retorne para saber como desbloquear.
Conclusão
Considerações Finais
Chegamos ao fim do estudo sobre os tecidos cartilaginoso, ósseo e muscular. Inicialmente aprendemos as características histológicas básicas dos tipos de cartilagem e onde podemos encontrá-los. Além disso, discutimos a formação do tecido cartilaginoso e os dois tipos de crescimento da cartilagem. Em seguida, iniciamos um interessante estudo sobre o tecido ósseo: vimos seus componentes principais, a estrutura básica dos ossos imaturos e maduros e os processos de ossificação. Ainda nesse conteúdo, também debatemos a importância e os tipos de articulações.
Por fim, estudamos o terceiro tipo dos tecidos básicos do corpo: o tecido muscular. Debatemos as características gerais e as funções dos músculos, bem como nos aprofundamos nos aspectos celulares e histológicos dos músculos estriados (esquelético e cardíaco) e lisos. Os mecanismos moleculares do processo de contração muscular também foram abordados com detalhes.
Foi uma longa jornada, repleta de aprendizados e conceitos novos que serão extremamente úteis para a sua carreira. Tais conceitos se aplicam a diversos campos, como Anatomia, Fisiologia, Patologia e Biologia celular.
Podcast
Agora, a especialista Gabriela Cardoso Caldas encerra com uma entrevista com a conteudista abordando os principais assuntos do tema.
CONQUISTAS
Você atingiu os seguintes objetivos:
Descreveu as características morfológicas, a função e a localização do tecido cartilaginoso, bem como os principais tipos de cartilagem e as suas particularidades
Descreveu as características morfológicas do tecido ósseo, os componentes fundamentais, a estrutura geral e a formação dos diferentes tipos de ossos
Reconheceu os três tipos de tecido muscular, as suas características morfológicas e as diferentes formas de contração muscular